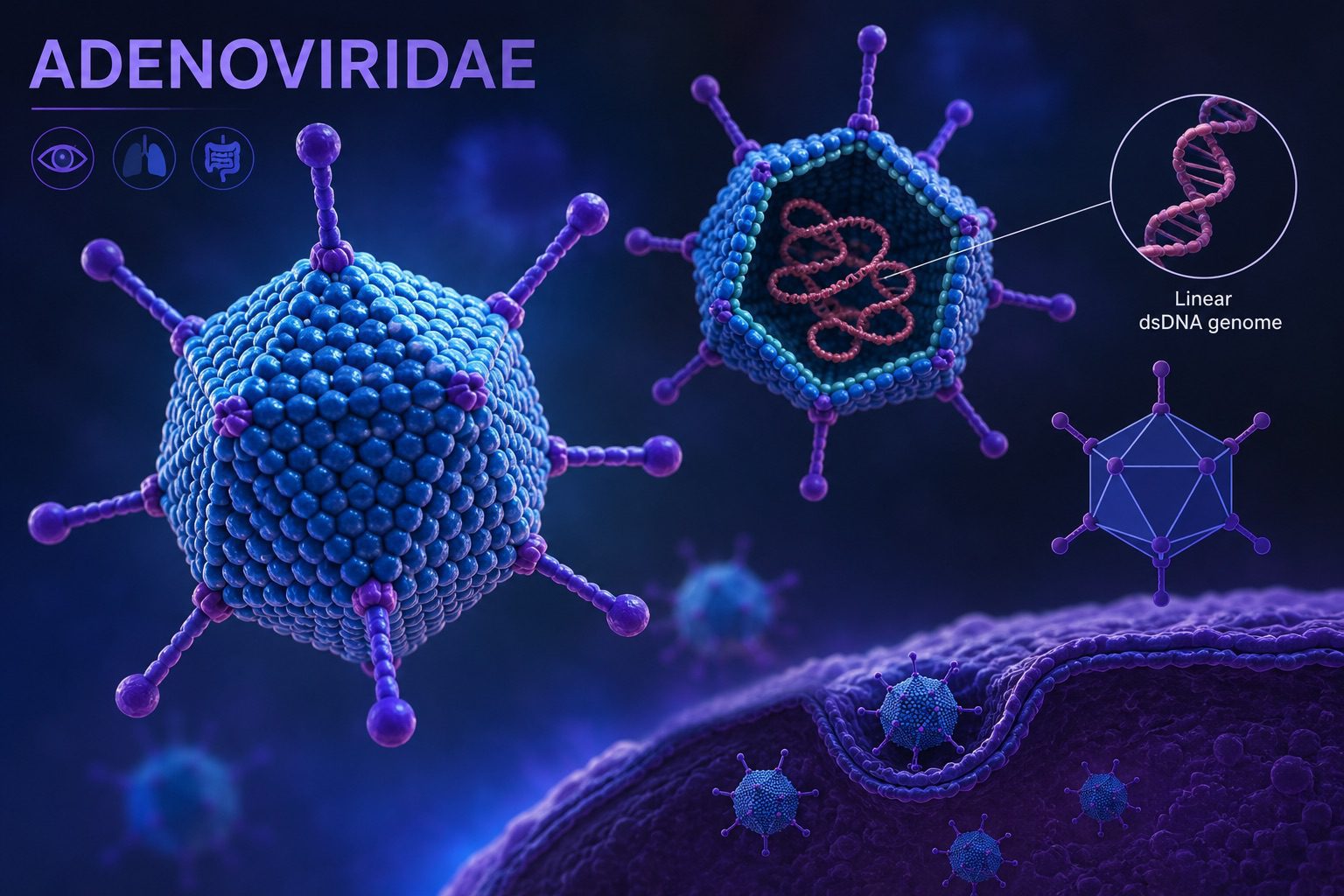
Viral composition refers to the collective assemblage of viral entities present within a defined environment, encompassing their taxonomic diversity, genome types, structural features, and ecological roles. These communities are extraordinarily heterogeneous, reflecting the immense evolutionary plasticity of viral populations and their intimate coupling with host organisms across all biological domains. Unlike cellular microbiota, viral assemblages are not constrained by a universal phylogenetic marker, which makes compositional assessment dependent on metagenomic inference, signature gene profiling, and de novo assembly strategies. Consequently, viral composition is often described in terms of operational viral units and genome-resolved clusters rather than strict species boundaries.
Across ecosystems, viral communities are typically dominated by bacteriophages, particularly within microbial-rich habitats such as soils, aquatic systems, and animal-associated niches. These phages exhibit a wide range of morphologies and replication strategies, including lytic and temperate lifestyles, which together shape microbial population dynamics and gene flow. In marine environments, for instance, double-stranded DNA viruses frequently dominate, especially members of Caudoviricetes-like morphotypes, while single-stranded DNA viruses can contribute significantly to background diversity. RNA viruses, although often less abundant in metagenomic datasets due to methodological biases, can represent highly dynamic components of viral assemblages, particularly in eukaryote-rich environments.
Host-associated viral communities, such as those found in gastrointestinal tracts, respiratory systems, or plant rhizospheres, often display strong co-structuring with host physiology and immune status. These viromes are typically enriched in prophages integrated within bacterial genomes, reflecting long-term co-evolutionary relationships. The balance between lytic replication and lysogenic integration is a key determinant of compositional stability, with environmental perturbations such as antibiotic exposure, diet shifts, or temperature fluctuations frequently triggering prophage induction and consequent restructuring of viral populations.
In soil ecosystems, viral composition is shaped by extreme spatial heterogeneity and physicochemical complexity. Soil viromes often contain a high proportion of novel viral lineages, many of which lack homologs in reference databases. This “viral dark matter” reflects both the under-sampling of terrestrial systems and the rapid diversification of viral genomes under selective pressures imposed by dense microbial consortia. Gene exchange mediated by viruses, particularly through transduction, contributes significantly to horizontal gene transfer, including the dissemination of metabolic and resistance-related genes.
Aquatic viral communities are among the most abundant biological entities on Earth, with concentrations often exceeding 107 particles per milliliter in marine systems. Their composition is strongly influenced by nutrient availability, host abundance, and water column stratification. Viral shunt processes, whereby infection-induced cell lysis redirects organic carbon from higher trophic levels into dissolved organic matter pools, further underscore the ecological importance of viral community structure. Seasonal blooms of microbial hosts often coincide with shifts in viral diversity, producing transient but highly structured compositional patterns.
From a genomic perspective, viral composition is characterized by mosaicism, high mutation rates, and frequent recombination events. These features generate extensive intra-population diversity, often referred to as quasispeciesdynamics in RNA viruses. DNA viruses, while generally more genetically stable, still exhibit significant modular evolution driven by horizontal gene exchange and genome rearrangements. Functional annotation of viral communities reveals enrichment in auxiliary metabolic genes, which can modulate host metabolism during infection and thereby influence ecosystem-level biogeochemical cycles.
Viral composition is not static but highly responsive to environmental conditions, host community structure, and evolutionary pressures. Its characterization requires integrative approaches combining metagenomics, viromics, and computational ecology. Understanding this complexity provides critical insights into microbial regulation, ecosystem functioning, and the evolutionary trajectories of both viruses and their hosts.
Structural architecture and molecular composition of virions
Viruses are infectious biological agents characterized by a fundamentally acellular organization and a highly streamlined structural design. At their core, virions consist of a nucleic acid genome enclosed within a proteinaceous capsid. The genome may be composed of either deoxyribonucleic acid (DNA) or ribonucleic acid (RNA), and it encodes the essential information required for viral replication, assembly, and host manipulation. This minimalist architecture distinguishes viruses from cellular life forms and underpins their obligate dependence on host cellular machinery for propagation.
The capsid, formed from repeating protein subunits known as capsomers, serves multiple critical functions. It provides physical protection to the viral genome against environmental degradation, facilitates attachment to susceptible host cells, and contributes to genome delivery during the early stages of infection. Capsid proteins are often highly organized into symmetrical structures, such as icosahedral or helical forms, reflecting evolutionary optimization for stability and efficient self-assembly.
In addition to this core structure, many viruses possess a lipid-containing outer layer known as an envelope. Enveloped viruses acquire this membrane from host cellular membranes during the budding process. Embedded within the envelope are viral glycoproteins that project outward as spikes, playing a central role in host receptor recognition, membrane fusion, and immune evasion. These glycoproteins are among the most functionally significant viral components, as they determine host range and tissue tropism.
In contrast, viruses that lack this lipid layer are referred to as naked viruses. These viruses rely solely on their capsid for environmental resilience and host interaction, often exhibiting greater resistance to desiccation, chemical agents, and physical stress compared to enveloped counterparts. The absence of an envelope also influences their transmission routes and stability outside host organisms.
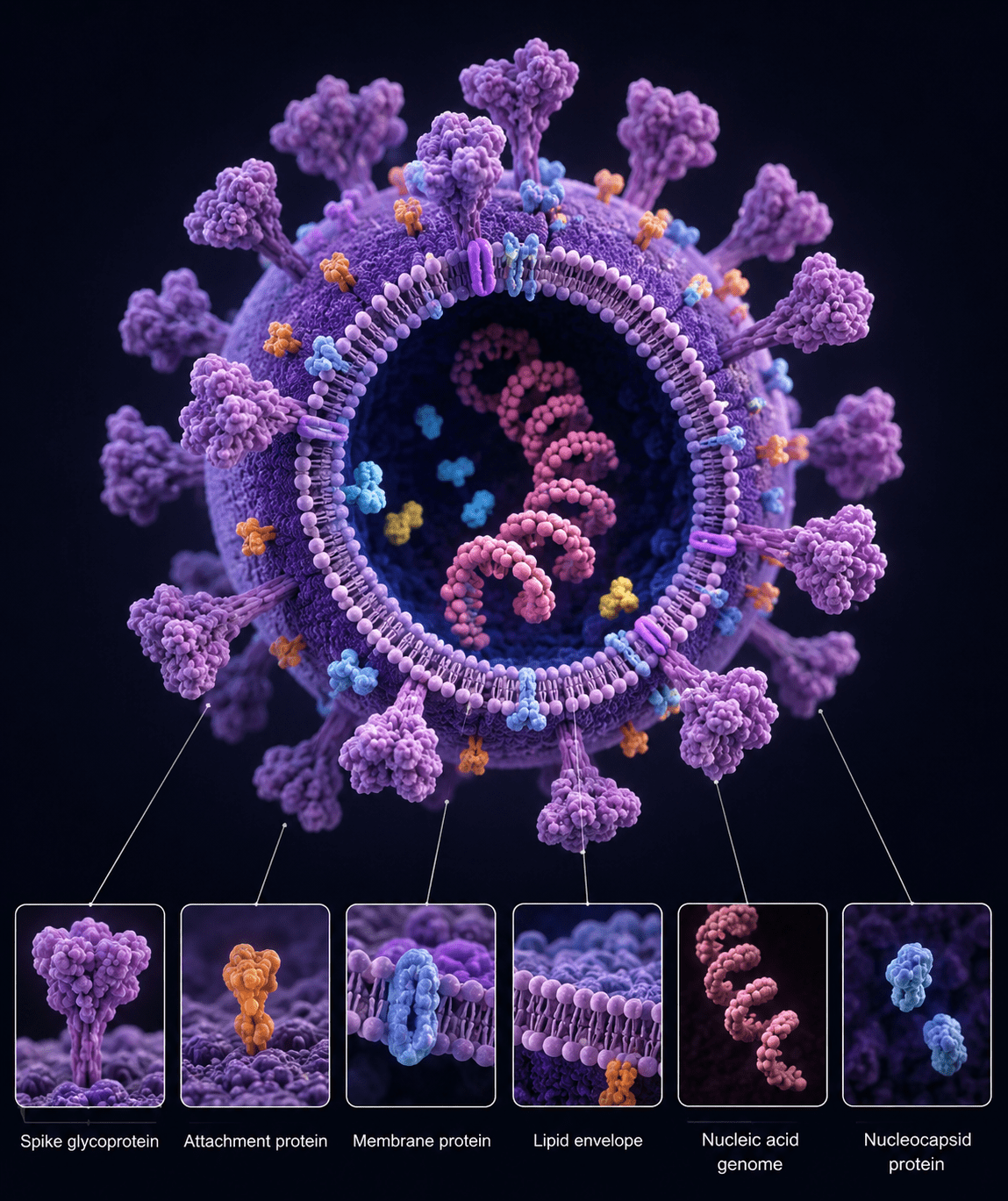
Beyond these structural elements, virions contain a coordinated set of molecular components that are essential for infectivity (Figure 1). Viral proteins include not only structural constituents of the capsid and envelope but also enzymatic and regulatory proteins that facilitate genome replication and transcription. Some viruses package enzymes such as polymerases directly within the virion to enable immediate transcription upon entry into the host cell. Viral genomes, whether segmented or non-segmented, single-stranded or double-stranded, further contribute to diversity in replication strategies and evolutionary dynamics. The chemical composition of viruses represents a finely tuned system optimized for survival, transmission, and replication within specific host environments. This structural and molecular economy is central to viral success across biological systems.
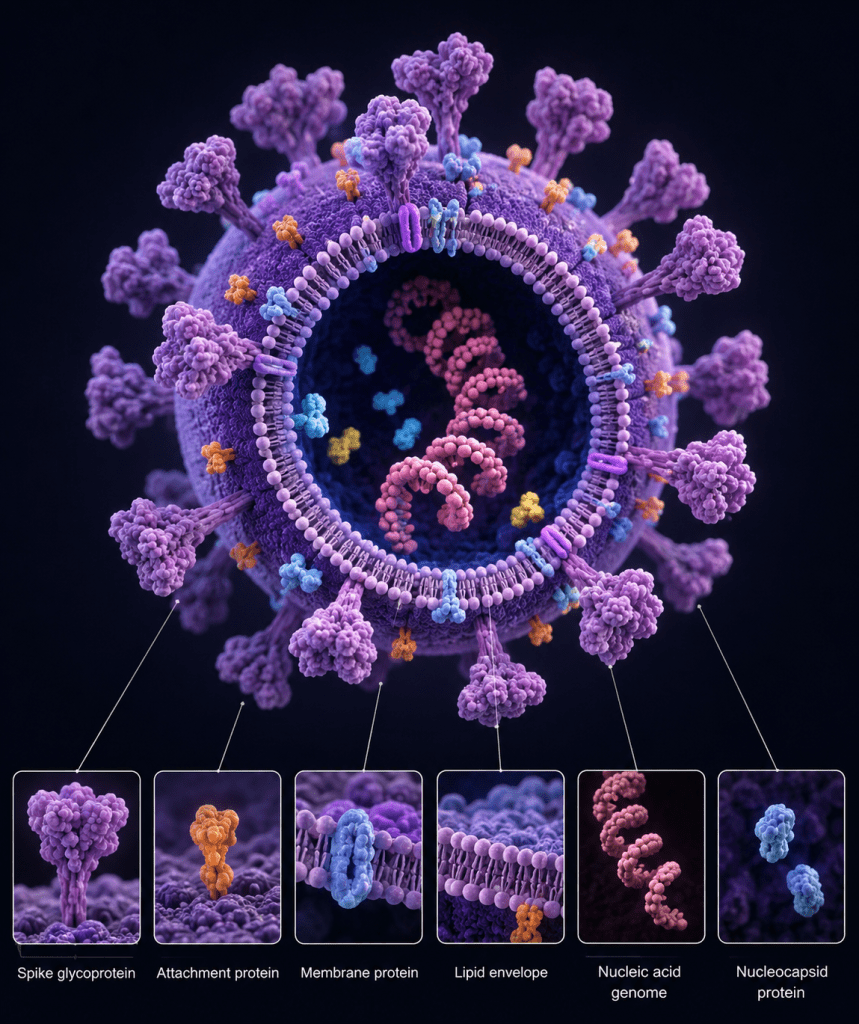
Viral Components: Structure, Function, and Mechanisms of Host Exploitation
1. Viral proteins: functional diversity and biological roles of viral proteins in infection and replication
Viral proteins constitute the fundamental structural and functional framework of viruses, underpinning virtually every stage of the viral life cycle, from host recognition and entry to replication, assembly, and immune interaction. These proteins are essential determinants of viral infectivity and are central to the biological identity of each virus. Importantly, they serve as the primary antigenic determinants (epitopes) recognized by the host immune system, making them key targets of neutralizing antibodies and cell-mediated immune responses, particularly T lymphocytes. This antigenic specificity forms the basis of serological diagnostics and vaccine development, as protective immunity is typically directed against exposed viral protein structures, especially those located on the virion surface.
A primary and highly specialized function of viral proteins is the mediation of host cell attachment and entry. Surface-exposed viral proteins, commonly referred to as attachment proteins, glycoproteins, or spike proteins, interact with specific receptor molecules on the surface of susceptible host cells. This receptor-ligand interaction is highly selective and is a major determinant of host range, tissue tropism, and species specificity. Even minor structural variations in these proteins can significantly alter viral infectivity and cross-species transmission potential. Following receptor engagement, viral proteins facilitate entry through mechanisms such as membrane fusion, receptor-mediated endocytosis, or direct penetration, depending on viral classification.
In enveloped viruses, viral glycoproteins embedded within the lipid envelope undergo conformational rearrangements that drive fusion between the viral envelope and the host cell membrane, enabling release of the viral nucleocapsid into the host cytoplasm. In contrast, non-enveloped viruses rely on capsid-associated proteins to disrupt endosomal membranes or form translocation pores, ensuring genome delivery into the host cellular environment.
Structurally, viral proteins are the primary constituents of the capsid, also known as the protein coat, which encloses and protects the viral genome, whether composed of DNA or RNA. Capsid proteins self-assemble into highly ordered supramolecular structures exhibiting defined symmetry, most commonly icosahedral or helical configurations. This structural organization is not arbitrary; it reflects evolutionary optimization for genome packaging efficiency, mechanical stability, and environmental resilience. The capsid acts as a robust protective barrier, shielding the viral genome from physical stress, chemical degradation, and enzymatic attack, particularly from host nucleases that would otherwise rapidly degrade exposed nucleic acids.
In enveloped viruses, additional structural proteins, such as matrix or tegument proteins, provide critical internal scaffolding that connects the nucleocapsid to the lipid envelope, ensuring virion integrity and coordinating assembly during viral budding. These proteins also play roles in regulating virion morphology and stability during extracellular transmission. Beyond structural roles, a substantial proportion of viral proteins are non-structural and are expressed exclusively during infection. These proteins are indispensable for viral replication and include enzymes such as polymerases, proteases, helicases, and integrases. They orchestrate genome replication, transcriptional regulation, protein processing, and virion assembly. Furthermore, many viral proteins actively manipulate host cellular pathways, suppress innate immune signaling, inhibit antigen presentation, and redirect cellular resources toward viral production. Through these mechanisms, viruses enhance their replication efficiency while evading immune detection. Viral proteins integrate structural integrity with dynamic regulatory functions, making them indispensable to viral survival, pathogenicity, and transmission. Their multifunctionality reflects a highly optimized evolutionary strategy that enables viruses to exploit host systems with remarkable efficiency and precision.
2. Viral enzymes: catalytic drivers of genome replication and host reprogramming
Viral enzymes are specialized catalytic proteins encoded by viral genomes that enable and regulate essential stages of the viral life cycle within host cells. Although viruses are obligate intracellular parasites that depend extensively on host metabolic and translational machinery for energy production and biosynthesis, many have evolved to encode or package a distinct set of enzymes that ensure successful genome replication, transcription, integration, and maturation. These enzymes are proteinaceous in nature and may be incorporated into the virion during assembly or synthesized immediately following host cell entry, depending on the viral replication strategy.
A central example of such enzymatic machinery is reverse transcriptase (RT), an RNA-dependent DNA polymerase characteristic of retroviruses such as Human immunodeficiency virus type 1. Reverse transcriptase catalyzes the synthesis of complementary DNA (cDNA) from an RNA template, thereby converting the viral RNA genome into a DNA intermediate. This step is fundamental because it enables the stable integration of viral genetic material into the host genome. The integrated viral DNA, termed a provirus, becomes a persistent genetic element within host chromosomes. This integration process is mediated by a second critical viral enzyme, integrase, which facilitates strand transfer and stable insertion of viral DNA into host chromatin. Once integrated, the provirus exploits host RNA polymerase II and associated transcriptional machinery to direct the synthesis of viral mRNAs and genomic RNA, effectively hijacking cellular gene expression systems. In the absence of reverse transcriptase, retroviruses would be incapable of establishing productive infection cycles.
Similarly, a wide range of RNA viruses encode RNA-dependent RNA polymerase (RdRp), an enzyme that synthesizes RNA from an RNA template. This enzymatic function is indispensable because eukaryotic and prokaryotic host cells do not possess endogenous polymerases capable of replicating RNA genomes. For instance, severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) relies on RdRp to replicate its positive-sense single-stranded RNA genome and to generate a nested set of subgenomic RNAs. These subgenomic transcripts are essential for the translation of structural and accessory proteins required for virion assembly and host modulation. RdRp activity is therefore central not only to genome amplification but also to the regulation of viral gene expression dynamics.
In addition to polymerases and integrases, many viruses encode auxiliary enzymes such as proteases, helicases, and capping enzymes. Viral proteases are responsible for processing large polyprotein precursors into functional units, a strategy commonly observed in positive-sense RNA viruses. Helicases facilitate the unwinding of nucleic acid structures during replication, while capping enzymes modify viral RNA to enhance stability and evade host immune recognition. Viral enzymes are generally non-structural components, meaning they do not contribute to the architecture of the capsid or envelope. Instead, they perform highly specialized catalytic roles that drive temporal regulation of infection, replication fidelity, and host reprogramming. Because of their essentiality and virus-specific nature, viral enzymes represent major targets for antiviral therapeutics, as inhibition of their activity can effectively block viral propagation without directly harming host cellular processes.
3. Viral genomes: architecture, diversity, and replication strategies of viral genomes
The viral genome represents the core repository of genetic information that directs viral replication, assembly, and propagation within a susceptible host cell. It encodes all functional determinants required for infection, including structural proteins, replication enzymes, and regulatory elements that modulate host-virus interactions. Unlike cellular organisms, which universally employ double-stranded DNA as their hereditary material, viruses display extraordinary heterogeneity in both the chemical nature and structural organization of their genomes. A foundational principle in virology is that a given virus contains either DNA or RNA as its genetic material, but never both simultaneously. This strict dichotomy underpins viral classification frameworks and exerts profound influence on replication mechanisms, evolutionary trajectories, and adaptability.
Viral genomes exhibit multiple architectural forms that contribute to their functional versatility. They may be composed of single-stranded (ss) or double-stranded (ds) nucleic acid, and may exist in either linear or circular configurations. These structural variations are not merely descriptive but are mechanistically significant, as they dictate how viral genomes are replicated, transcribed, and packaged into progeny virions. Double-stranded viral genomes often resemble cellular genetic material and can exploit host or virus-encoded DNA-dependent polymerases for replication and transcription. In contrast, single-stranded genomes require conversion into complementary strands before gene expression or genome duplication can occur, necessitating additional enzymatic steps and specialized replication strategies.
Another fundamental feature of viral genome organization is segmentation. Viral genomes may be segmented, in which the genetic information is distributed across multiple distinct nucleic acid molecules, or non-segmented, where the genome exists as a single continuous molecule. Segmentation introduces a powerful evolutionary mechanism known as reassortment, whereby genome segments are exchanged between related viruses co-infecting the same host cell. This process can generate abrupt genetic shifts and significantly enhance viral diversity, sometimes leading to emergent strains with altered virulence or host range. Non-segmented genomes, by contrast, evolve primarily through point mutations and homologous recombination, resulting in more gradual genetic change.
Within RNA viruses, genome polarity or sense constitutes a critical determinant of replication strategy. Positive-sense (+) single-stranded RNA viruses possess genomes that function directly as messenger RNA. Upon entry into the host cytoplasm, their genomic RNA can be immediately translated by host ribosomes to produce viral proteins, including those required for replication. Negative-sense (−) single-stranded RNA viruses, however, carry genomes that are complementary to mRNA and therefore cannot be directly translated. These viruses must package an RNA-dependent RNA polymerase within the virion to synthesize a positive-sense intermediate, which then serves as the template for protein synthesis. A further refinement of RNA genome organization is the ambisense strategy, in which coding sequences are present in both positive and negative orientations within the same genome segment. This arrangement necessitates temporally regulated transcription from both strands during infection and is characteristic of viral families such as Arenaviridae, highlighting the evolutionary flexibility of RNA genome architectures.
The biochemical and structural characteristics of viral genomes also vary significantly in size and complexity. DNA viruses generally possess larger genomes, typically ranging from approximately 3 to 370 kilobase pairs (kbp). These genomes benefit from relatively high replication fidelity, owing to the proofreading capabilities of many DNA polymerases, which contributes to their greater genetic stability. RNA viruses, in contrast, tend to have smaller genomes, usually between 7 and 30 kilobases (kb). This size limitation is largely imposed by the intrinsically higher error rates of RNA-dependent RNA polymerases, which lack efficient proofreading mechanisms. As a result, RNA viruses exhibit elevated mutation rates, fostering rapid evolution, antigenic drift, and the capacity to adapt swiftly to environmental pressures such as host immune responses and antiviral interventions.
In animal virology, several broad patterns of genome organization are consistently observed. The majority of viruses infecting animals and humans possess double-stranded DNA genomes, reflecting structural stability and compatibility with host nuclear processes. A notable exception is the family Parvoviridae, whose members contain single-stranded DNA genomes, requiring host or viral machinery to synthesize complementary strands prior to gene expression. Conversely, most RNA viruses infecting animals and humans are characterized by single-stranded RNA genomes. The principal exception to this rule is the family Reoviridae, which contains double-stranded RNA genomes that are replicated within specialized viral replication complexes to avoid host immune detection. The diversity in viral genome type, structure, segmentation, and polarity reflects a range of evolutionary solutions to the central biological challenge of efficient replication within host environments while evading immune defenses. Genome architecture governs not only replication strategy but also pathogenic potential, host specificity, transmission dynamics, and mechanisms of genetic variation.
4. Viral glycoproteins: structure, biosynthesis, and functional roles in infection
Viral glycoproteins are highly specialized macromolecules consisting of a polypeptide backbone covalently linked to carbohydrate chains through enzymatic glycosylation. These proteins are encoded by viral genomes but are synthesized and extensively processed within infected host cells, relying heavily on the host’s translational apparatus and secretory pathway. Following translation on ribosomes associated with the rough endoplasmic reticulum (ER), nascent polypeptides undergo folding, disulfide bond formation, and co- and post-translational glycosylation. Glycan addition occurs primarily in the ER and is subsequently modified in the Golgi apparatus, where complex oligosaccharide structures are processed and refined. These modifications are typically mediated through N-linked glycosylation, involving attachment to asparagine residues, or O-linked glycosylation, involving serine or threonine residues.
Once properly folded and glycosylated, viral glycoproteins are trafficked through the host secretory system and targeted to specific cellular membranes. In enveloped viruses, these proteins are inserted into lipid bilayers derived from host cell membranes during viral assembly and budding. The resulting virions therefore display glycoprotein spikes embedded within their external envelope, forming a critical interface between virus and host.
Functionally, viral glycoproteins are central to the infectious cycle. They mediate the initial attachment of virions to host cell receptors, a process known as adsorption. This interaction is highly specific and is a primary determinant of host range, tissue tropism, and species specificity. For example, the hemagglutinin (HA) and neuraminidase (NA) glycoproteins of Influenza A virus coordinate receptor binding, membrane fusion, and viral release. Similarly, the spike (S) glycoprotein of SARS-CoV-2 binds to the angiotensin-converting enzyme 2 (ACE2) receptor and facilitates fusion of viral and cellular membranes.
Beyond receptor recognition, many viral glycoproteins actively mediate membrane fusion, enabling entry of the viral genome into the host cytoplasm. This fusion process often involves substantial conformational rearrangements triggered by receptor binding or endosomal acidification, allowing the viral envelope to merge with host membranes. In this way, glycoproteins function not only as attachment factors but also as molecular machines that drive viral entry.
In addition to their mechanistic roles, viral glycoproteins are among the most immunologically significant components of virions. As surface-exposed antigens, they are primary targets of neutralizing antibodies generated by the host immune system. Consequently, they play a central role in protective immunity and are key focal points in vaccine development and antiviral drug design. However, many viruses exploit glycosylation as a strategy for immune evasion. Dense glycan shields can obscure conserved protein epitopes, reducing antibody accessibility and facilitating persistent infection.
In contrast, non-enveloped or “naked” viruses lack lipid membranes and therefore do not incorporate glycoproteins into an external envelope. Instead, they rely on structural capsid proteins to mediate receptor binding and cell entry, often through different mechanistic pathways. Viral glycoproteins represent multifunctional molecular interfaces that integrate structural biology, host-cell exploitation, and immune system interaction, making them indispensable determinants of viral infectivity and pathogenesis.
5. Viral envelopes: structural origin, composition, and functional significance in viral infectivity
Viruses are broadly categorized into enveloped and non-enveloped (naked) forms based on the presence or absence of an external lipid membrane. This distinction is not merely structural but has profound consequences for viral infectivity, environmental persistence, transmission pathways, and susceptibility to physical and chemical inactivation. The viral envelope represents one of the most important adaptations in viral evolution, particularly among viruses that rely on efficient host cell entry and immune evasion.
Enveloped viruses possess an outer lipid bilayer termed the viral envelope. This membrane is not independently synthesized by the virus; instead, it is acquired from the host cell during the late stages of viral replication and assembly. After viral genome replication and capsid formation within the host cell, newly assembled nucleocapsids are transported to specific cellular membranes. Depending on the virus, these membranes may include the plasma membrane, nuclear membrane, endoplasmic reticulum, or Golgi apparatus. The selection of the membrane source is virus-specific and often reflects the intracellular site of viral assembly.
The acquisition of the envelope occurs through a process known as budding. During budding, the nucleocapsid pushes outward through the host-derived lipid bilayer, incorporating a segment of the membrane into its final structure. This process allows the virus to exit the host cell without immediate lysis in many cases, thereby facilitating persistent infection and prolonged viral shedding. Embedded within the lipid envelope are viral glycoproteins, which are encoded by the viral genome and selectively inserted into the host membrane prior to or during budding. These glycoproteins, often appearing as spike-like projections on the virion surface, are essential for mediating attachment to host cell receptors, determining host range (tropism), and driving membrane fusion during entry.
Because the envelope is composed primarily of lipids, enveloped viruses are structurally fragile in extracellular environments. Lipid bilayers are inherently sensitive to disruption by physical and chemical agents. Consequently, enveloped viruses are highly susceptible to desiccation, elevated temperatures, detergents, alcohol-based disinfectants, ether, and other lipid-solvent compounds. Disruption of the envelope leads to loss of glycoprotein function, preventing receptor binding and fusion, and thereby rendering the virus non-infectious. This fragility explains why enveloped viruses typically require relatively direct transmission routes, such as respiratory droplets, saliva, blood, or other bodily fluids, and are less stable on environmental surfaces.
In contrast, non-enveloped (naked) viruses lack this lipid membrane and consist solely of a nucleic acid genome enclosed within a robust protein capsid. The absence of a lipid envelope confers significantly greater structural stability and environmental resilience. Naked viruses are generally resistant to detergents, alcohols, and many disinfectants that target lipid membranes. They also tolerate broader ranges of temperature, pH variation, and desiccation. As a result, they can persist for extended periods outside a host organism, increasing their likelihood of indirect transmission through contaminated surfaces (fomites), water, or food, particularly via the fecal-oral route. The presence or absence of a viral envelope is a defining structural feature that shapes nearly every aspect of viral biology, from environmental survival and transmission efficiency to host interaction and inactivation susceptibility.
References
Acheson N.H (2011). Fundamentals of Molecular Virology. Second edition. John Wiley and Sons Limited, West Sussex, United Kingdom.
Alan J. Cann (2005). Principles of Molecular Virology. 4th edition. Elsevier Academic Press, Burlington, MA, USA.
Alberts B, Bray D, Johnson A, Lewis J, Raff M, Roberts K and Walter P (1998). Essential Cell Biology: An Introduction to the Molecular Biology of the Cell. Third edition. Garland Publishing Inc., New York.
Barrett J.T (1998). Microbiology and Immunology Concepts. Philadelphia, PA: Lippincott-Raven Publishers. USA.
Black, J.G. (2008). Microbiology: Principles and Explorations (7th ed.). Hoboken, NJ: J. Wiley & Sons.
Brian W.J Mahy and Mark H.C van Regenmortel (2010). Desk Encyclopedia of Human and Medical Virology. Elsevier Academic Press, San Diego, USA.
Brooks G.F., Butel J.S and Morse S.A (2004). Medical Microbiology, 23rd edition. McGraw Hill Publishers. USA.
Cann A.J (2011). Principles of Molecular Virology. Fifth edition. Academic Press, San Diego, United States.
Carter J and Saunders V (2013). Virology: Principles and Applications. Second edition. Wiley-Blackwell, New Jersey, United States.
Champoux J.J, Neidhardt F.C, Drew W.L and Plorde J.J (2004). Sherris Medical Microbiology: An Introduction to Infectious Diseases. 4th edition. McGraw Hill Companies Inc, USA.
Dimmock N (2015). Introduction to Modern Virology. Seventh edition. Wiley-Blackwell, New Jersey, United States.
Dimmock N.J, Easton A.J and Leppard K.N (2001). Introduction to modern virology. 5th edition. Blackwell Science publishers. Oxford, UK.
Discover more from Microbiology Class
Subscribe to get the latest posts sent to your email.