Virus morphology refers to the size, shape, and structural organization of viruses. Although viruses are extremely small and can only replicate inside living host cells, they display remarkable structural diversity that influences how they infect hosts, survive in the environment, and spread between organisms. A typical virus consists of genetic material enclosed within a protective protein coat. The genetic material may be either DNA or RNA and can exist as single-stranded or double-stranded forms. Surrounding this genetic core is the capsid, which is composed of protein subunits called capsomeres. Together, the nucleic acid and capsid form the nucleocapsid, the fundamental structural unit of a virus. Some viruses possess an additional outer membrane known as the envelope, which is derived from the host cell membrane during viral release. Enveloped viruses contain embedded glycoprotein spikes that facilitate attachment and entry into host cells. In contrast, non-enveloped (naked) viruses lack this membrane and are generally more resistant to environmental conditions.
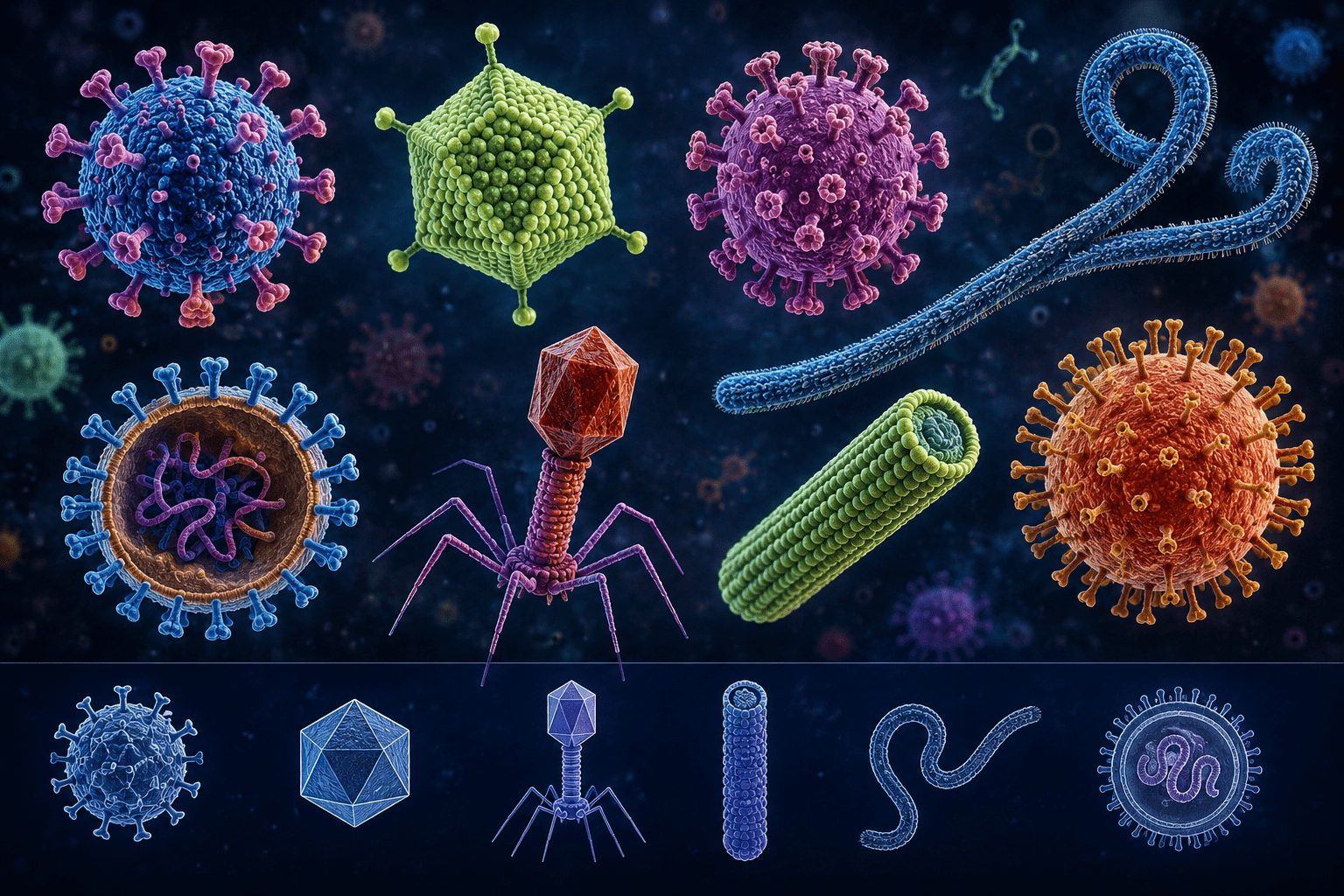
Viruses are generally classified into several morphological categories. Helical viruses possess capsomeres arranged in a spiral around the nucleic acid, creating a rod-like or filamentous appearance. This morphology is common among many plant viruses and certain animal viruses. Icosahedral viruses exhibit a symmetrical structure with twenty triangular faces, providing stability and efficient packaging of genetic material. Numerous human and animal viruses display this geometric arrangement. A further category includes complex viruses, which exhibit structures that do not fit purely into helical or icosahedral patterns. Certain bacteriophages, for example, possess a polyhedral head attached to a tail structure used for injecting genetic material into bacterial cells. Viral morphology is closely linked to viral function, host specificity, transmission, and pathogenicity.
A complete, infectious viral particle is known as a virion. Although viruses are acellular and lack independent metabolic machinery, they possess highly organized structural components that enable them to infect host cells efficiently. Despite their simplicity, viruses exhibit remarkable diversity in shape, symmetry, genome type, and structural complexity. The morphology of viruses reflects an elegant balance between structural simplicity and functional sophistication. Despite lacking cellular organization, viruses possess highly ordered architectures optimized for genome protection, host attachment, and replication. Understanding viral morphology is fundamental in virology because structure determines how viruses attach to host cells, penetrate tissues, replicate, and evade immune defenses. The structural design of a virus also plays a critical role in viral classification, pathogenesis, vaccine development, antiviral drug targeting, and diagnostic identification.
A comprehensive understanding of viral morphology is essential in virology, epidemiology, immunology, and biotechnology. Structural insights continue to inform antiviral drug design, vaccine development, and the study of viral evolution. Viral morphology provides the architectural blueprint that underlies infectivity, transmission, and pathogenic potential – making it a foundational concept in the study of viruses.
Essential elements governing viral structure, survival, and infectivity
A virion is the complete, mature, and infectious form of a virus outside the host cell. Despite their simple organization, virions possess highly specialized structural components that enable them to protect genetic material, recognize host cells, and initiate infection. A typical virion consists of three principal structural elements:
- The nucleic acid genome (DNA or RNA)
- The capsid (protein coat)
- The envelope (present in some viruses)
Each of these components contributes uniquely to viral infectivity, transmission, and survival. The nucleic acid genome serves as the genetic blueprint of the virus. Viral genomes may consist of either deoxyribonucleic acid (DNA) or ribonucleic acid (RNA) and may occur as single-stranded or double-stranded molecules depending on the virus type. This genetic material contains the information required for replication, synthesis of viral proteins, and establishment of infection within the host.
Surrounding the genome is the capsid, a protective protein shell composed of repeating protein subunits called capsomeres. The capsid protects viral nucleic acid from environmental damage and facilitates attachment and delivery into host cells. The arrangement of capsomeres determines viral shape and contributes to host specificity. Some viruses possess an additional envelope, a lipid membrane derived from the host cell during viral release. Embedded viral glycoproteins on the envelope mediate host-cell recognition, attachment, and entry, enhancing infectivity and immune interactions.
1. Viral nucleic acid genome: the genetic blueprint governing viral replication and infectivity
The viral nucleic acid genome is the fundamental hereditary component of a virus and serves as the molecular blueprint that directs viral replication, protein synthesis, and infection of host cells. Unlike cellular organisms, which contain both DNA and RNA as part of their biological systems, viruses possess only one type of nucleic acid in their virion structure either deoxyribonucleic acid (DNA) or ribonucleic acid (RNA), but never both simultaneously. This distinction is one of the defining characteristics of viruses.
Viral genome sizes vary considerably, ranging from approximately 3 kilobases (kb) in small RNA viruses to more than 300 kilobases (kb) in large DNA viruses, reflecting the broad diversity and adaptive capacity of viruses. Viral genomes exhibit remarkable diversity in composition and organization. Based on molecular structure, viral genomes can occur in several forms:
- Double-stranded DNA (dsDNA)
- Single-stranded DNA (ssDNA)
- Double-stranded RNA (dsRNA)
- Single-stranded RNA (ssRNA)
Beyond these structural categories, viral genomes differ in genome polarity, segmentation, and topology. RNA viruses may possess positive-sense (+) RNA, which can function directly as messenger RNA for protein synthesis, or negative-sense (−) RNA, which requires transcription into a complementary strand before translation. Some viruses have segmented genomes, where genetic information is divided into separate nucleic acid molecules, while others possess non-segmented genomes. Additionally, viral genomes may be arranged in linear or circular configurations, influencing replication mechanisms and genome stability. For example, viruses belonging to the family Adenoviridae contain double-stranded DNA genomes enclosed within an icosahedral capsid. In contrast, members of the family Orthomyxoviridae possess segmented single-stranded RNA genomes, a feature that facilitates genetic reassortment and contributes to viral evolution.
Beyond storing genetic information, the viral genome encodes proteins that regulate multiple essential biological functions. The viral genome performs functions beyond genetic storage. It encodes proteins essential for:
- Viral replication and transcription: The viral genome directs the synthesis of new viral nucleic acid molecules and controls the production of viral messenger RNA (mRNA). Replication ensures the generation of additional genome copies that will be packaged into progeny virions, while transcription produces mRNA templates for protein synthesis. Different virus groups employ distinct replication strategies depending on genome type; for example, RNA viruses often rely on specialized viral polymerases, whereas many DNA viruses use host-cell transcriptional machinery. Efficient replication and transcription are fundamental for sustaining infection and increasing viral load.
- Host-cell recognition and attachment: The viral genome encodes proteins involved in identifying and binding to specific receptors on host-cell surfaces. These receptor-binding proteins determine host range, tissue tropism, and the overall ability of a virus to establish infection. Viral surface proteins interact with complementary molecules on susceptible cells, initiating attachment and subsequent entry processes. This highly selective recognition mechanism explains why many viruses infect only particular species, tissues, or cell types and is a major determinant of viral pathogenicity.
- Immune evasion mechanisms: Many viral genomes encode proteins that enable viruses to avoid, suppress, or manipulate host immune responses. These mechanisms may include inhibition of antiviral signaling pathways, reduction of antigen presentation, interference with interferon activity, and alteration of host immune recognition. Some viruses establish persistent or latent infections by minimizing immune detection. Through these adaptations, viruses enhance their survival, prolong infection, and improve opportunities for transmission to new hosts.
- Structural assembly of new virions: The viral genome contains instructions for producing structural proteins required for constructing complete infectious particles. These proteins assemble into organized components such as the capsid, envelope-associated structures, and nucleoprotein complexes. During viral maturation, newly synthesized genomes are packaged into these structures through highly coordinated processes. Proper assembly ensures that progeny virions remain stable, infectious, and capable of initiating subsequent cycles of infection in susceptible host cells.
- Enzymatic activities, including polymerases and proteases: Many viruses encode enzymes essential for completing their replication cycle. Polymerases catalyze the synthesis of viral DNA or RNA and are particularly important in viruses that cannot depend entirely on host-cell enzymes. Proteases cleave large precursor proteins into functional units required for viral maturation and infectivity. Additional viral enzymes may facilitate genome integration, nucleic acid modification, or processing of viral components. These enzymatic functions are often targeted in antiviral drug development because they play indispensable roles in viral propagation.
2. The capsid: protective protein architecture and determinant of viral identity
The capsid is the proteinaceous outer shell that encloses and protects the viral nucleic acid genome. It is one of the most fundamental structural components of a virion and plays a central role in maintaining viral integrity, infectivity, and transmission. Unlike cellular organisms, viruses lack complex cellular membranes and organelles; therefore, the capsid serves as the primary structural framework that preserves the viral genome outside the host environment.
Capsids are composed of repeating protein subunits known as capsomeres, which assemble spontaneously through highly organized molecular interactions. This process of self-assembly enables efficient formation of stable viral particles while minimizing genetic requirements for structural complexity. The arrangement and number of capsomeres vary among viruses and contribute directly to viral morphology and biological behavior.
When the viral genome and capsid are considered together, they form the nucleocapsid, which represents the basic infectious unit of many viruses. The capsid of a virus performs several essential functions that extend beyond simple protection:
- Protection of the viral genome: One of the primary roles of the capsid is to shield viral genetic material from physical, chemical, and biological damage. Viral nucleic acids are vulnerable to degradation by environmental factors and host enzymes such as nucleases. The capsid provides a durable barrier that preserves genome stability during transmission between host cells and across external environments, thereby maintaining viral infectivity.
- Facilitation of host-cell attachment and entry: Capsid proteins frequently participate in the early stages of infection by interacting with specific receptors located on the surface of host cells. These interactions promote viral attachment and may initiate mechanisms of entry, including membrane fusion or endocytosis. In non-enveloped viruses particularly, the capsid serves as the primary structure responsible for mediating host-cell penetration.
- Determination of host specificity (tropism): The molecular characteristics of capsid proteins strongly influence viral tropism, which refers to the preference of a virus for particular host species, tissues, or cell types. Specific receptor-binding regions on the capsid determine whether infection can occur and contribute significantly to disease distribution and severity.
Provision of antigenic determinants: Capsid proteins contain molecular regions known as antigenic determinants (epitopes) that are recognized by the host immune system. These structures stimulate immune responses and are often targeted by antibodies. Consequently, capsid proteins are important in vaccine development, viral diagnostics, and immune surveillance.
The structural organization of capsid proteins gives rise to distinct viral morphologies. These arrangements include helical symmetry, icosahedral symmetry, and more complex structural forms, each reflecting adaptations that support efficient genome packaging, stability, and transmission. Thus, the capsid is not merely a protective covering but a multifunctional structure essential to the viral life cycle.
Classification of viruses based on capsid symmetry
Structural patterns of capsid organization and their role in viral morphology
From a morphological and structural standpoint, viruses are commonly classified according to the symmetry and spatial arrangement of proteins within the capsid. Capsid symmetry reflects how capsomeres are organized around the viral genome and determines the overall shape, stability, packaging efficiency, and infective properties of the virion. Because capsid architecture is closely associated with viral function and host interaction, it represents an important criterion in viral classification.
Based on capsid organization, viruses are broadly divided into three major structural symmetry groups:
- Icosahedral (cubic) symmetry: This group of viruses are characterized by a highly ordered geometric structure composed of twenty triangular faces that maximize internal volume while maintaining structural stability.
- Helical symmetry: This group of viruses are characterized by capsomeres arranged in a spiral or coiled pattern around the viral nucleic acid, producing rod-shaped or filamentous virions.
- Complex symmetry: This group of viruses are characterized by structural features that do not conform exclusively to either icosahedral or helical arrangements and often involve additional specialized components.
In addition to capsid symmetry, viruses may further be categorized as enveloped or non-enveloped, depending on the presence or absence of an outer lipid membrane surrounding the nucleocapsid. This additional feature significantly influences viral stability, transmission, and mechanisms of host-cell entry.
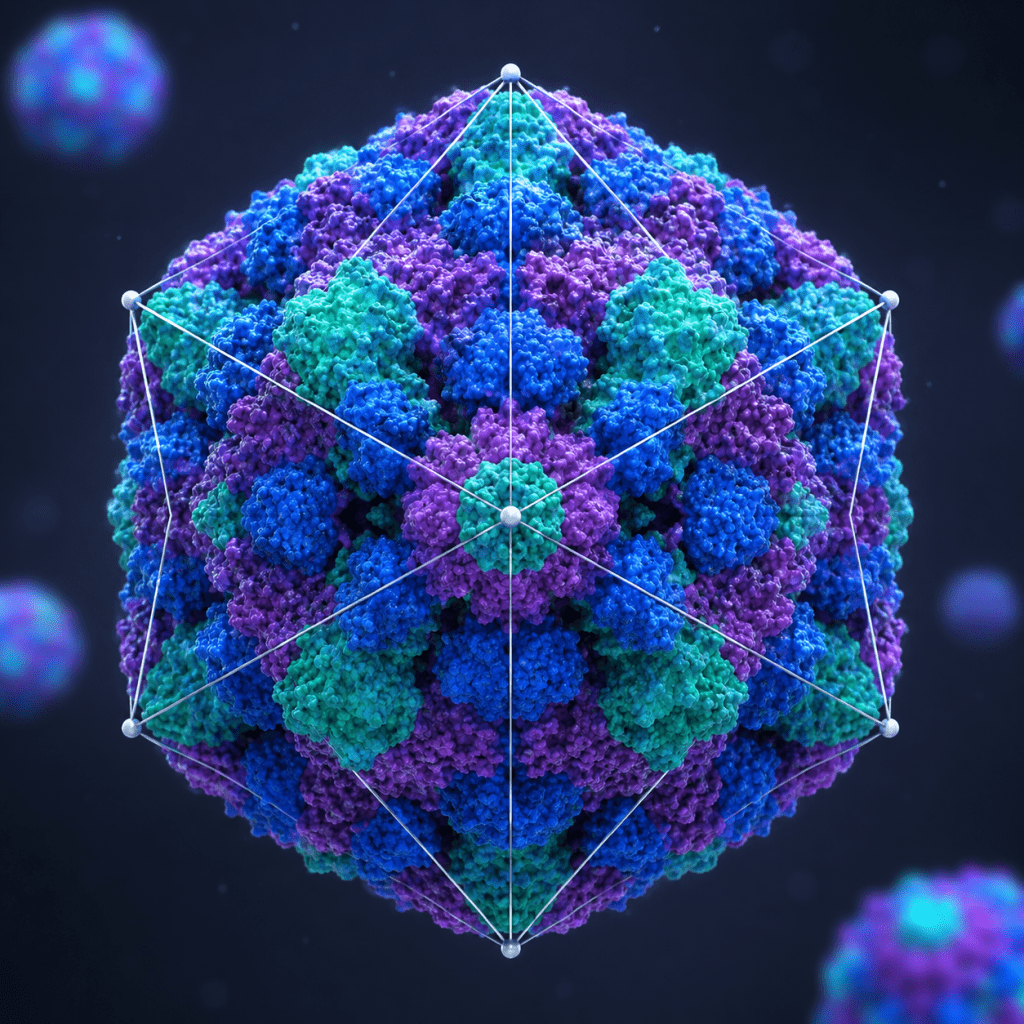
Icosahedral (cubic) viruses
Icosahedral viruses possess one of the most structurally efficient and widely distributed forms of viral architecture. Their capsid exhibits a highly symmetrical icosahedral (cubic) arrangement, consisting of 20 triangular faces, 12 vertices, and 30 edges that together create a closed protein shell surrounding the viral genome (Figure 1). This geometric design provides maximum internal volume with minimal structural protein requirements, making it an energetically favorable and evolutionarily successful form of viral organization.
The capsid is composed of repeating protein subunits called capsomeres, which self-assemble into a stable and ordered structure. This organization protects the viral nucleic acid while enabling efficient delivery of the genome into susceptible host cells. Although icosahedral viruses frequently appear spherical under electron microscopy, their true structure is a regular polyhedral form rather than a perfect sphere. Icosahedral symmetry occurs in both DNA and RNA viruses, demonstrating that this capsid arrangement is compatible with diverse genetic strategies. Viral genomes enclosed within these structures may be single-stranded or double-stranded and may occur with or without an external envelope.
An example of icosahedral virus family is the family Adenoviridae, whose members are non-enveloped icosahedral viruses associated with respiratory infections, conjunctivitis, and gastroenteritis. Representative viruses within this family include Human adenovirus A, Human adenovirus B, Human adenovirus C, and Human adenovirus F.
Other important icosahedral virus families and representative viruses include:
- Papillomaviridae
- Human papillomavirus (HPV-16, HPV-18)
- Picornaviridae
- Poliovirus
- Rhinovirus
- Hepatitis A virus
- Herpesviridae (enveloped with icosahedral nucleocapsid)
- Herpes simplex virus type 1 (HSV-1)
- Varicella-zoster virus (VZV)
- Epstein-Barr virus (EBV)
- Flaviviridae (enveloped with icosahedral nucleocapsid)
- Dengue virus
- Yellow fever virus
- Zika virus
Icosahedral viruses possess several important biological characteristics as follows:
- Efficient packaging of viral genomes
- High structural stability and protection of genetic material
- Strong antigenic properties due to exposed capsid proteins
- Adaptability to both enveloped and non-enveloped structural forms
Viruses with non-enveloped icosahedral capsids generally demonstrate greater resistance to environmental stressors such as drying, heat, acidity, and detergents, which enhances survival outside the host and facilitates transmission. Thus, the icosahedral capsid represents a highly optimized structural solution that supports viral persistence, infectivity, and evolutionary success.
Spherical symmetry in viruses
Some viruses exhibit spherical symmetry, a morphological appearance that is commonly observed under electron microscopy (Figure 2). This spherical shape is not always a true geometric symmetry in the strict structural sense but rather results from the combination of an underlying capsid arrangement and, in many cases, the presence of a surrounding lipid envelope. The envelope, derived from the host cell membrane, smooths the overall outline of the virion, giving it a rounded or spherical appearance.
Spherical viruses are typically enveloped viruses, meaning they possess an outer lipid bilayer embedded with viral glycoproteins. These glycoproteins play a crucial role in host-cell recognition, attachment, and membrane fusion, thereby facilitating viral entry. The internal structure of spherical viruses is often based on either icosahedral nucleocapsids or more complex arrangements that are masked by the envelope.
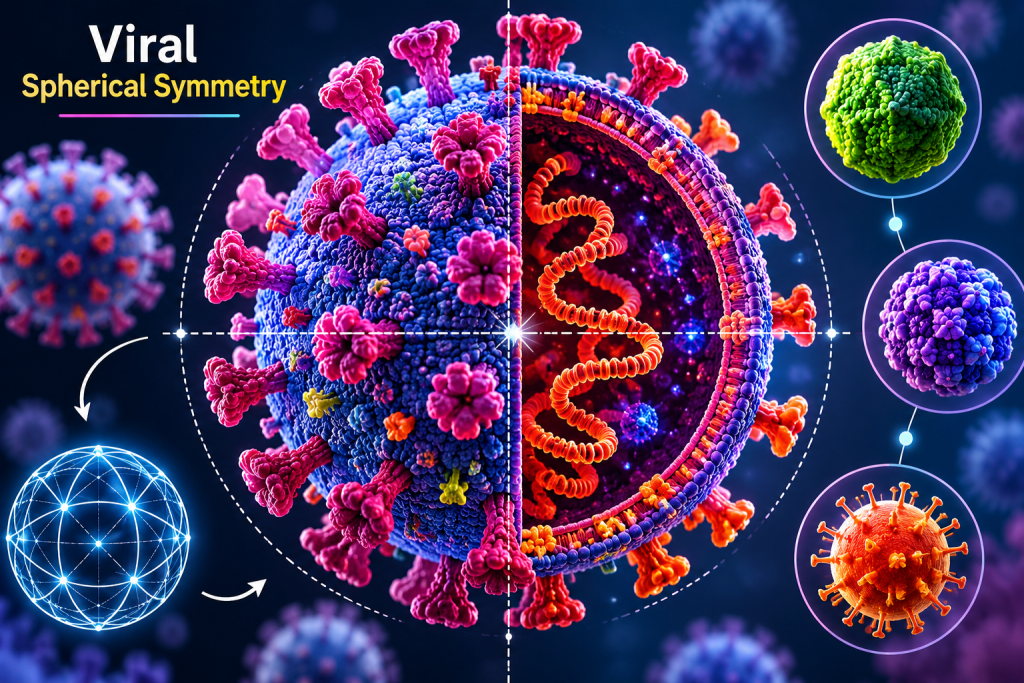
A wide range of medically important viruses exhibit spherical morphology. Examples include members of the families:
- Orthomyxoviridae such as Influenza A virus, which causes seasonal influenza epidemics.
- Coronaviridae such as the SARS-CoV-2, responsible for COVID-19.
- Retroviridae such as Human immunodeficiency virus (HIV). HIV also display spherical enveloped structures.
These viruses are generally more sensitive to environmental conditions compared to non-enveloped viruses, as the lipid envelope is easily disrupted by heat, detergents, and desiccation. However, their structural flexibility and surface glycoproteins enhance infectivity and immune interaction, making spherical viruses highly efficient in host invasion and transmission.
Helical viruses
Helical viruses are characterized by a distinctive structural organization in which capsid proteins are arranged in a continuous spiral (helical) configuration around the viral nucleic acid genome (Figure 3). In this architecture, the viral genome interacts with repeating protein subunits, known as capsomeres, which assemble in a regular helical pattern. This produces a highly ordered nucleocapsid with a rod-like, filamentous, or flexible thread-like appearance depending on the virus type.
A key structural feature of helical viruses is that their length is directly proportional to the size of the genome, while the diameter remains relatively constant. This is because capsomeres bind sequentially along the nucleic acid strand, extending the virion as the genome length increases. This structural principle allows helical viruses to efficiently accommodate varying genome sizes without altering the fundamental diameter of the capsid.
Helical nucleocapsids may exist in both rigid and flexible forms, depending on the strength of protein–protein and protein–nucleic acid interactions. In plant viruses, helical structures are often rigid, whereas many animal viruses display more flexible configurations due to the presence of an external envelope.
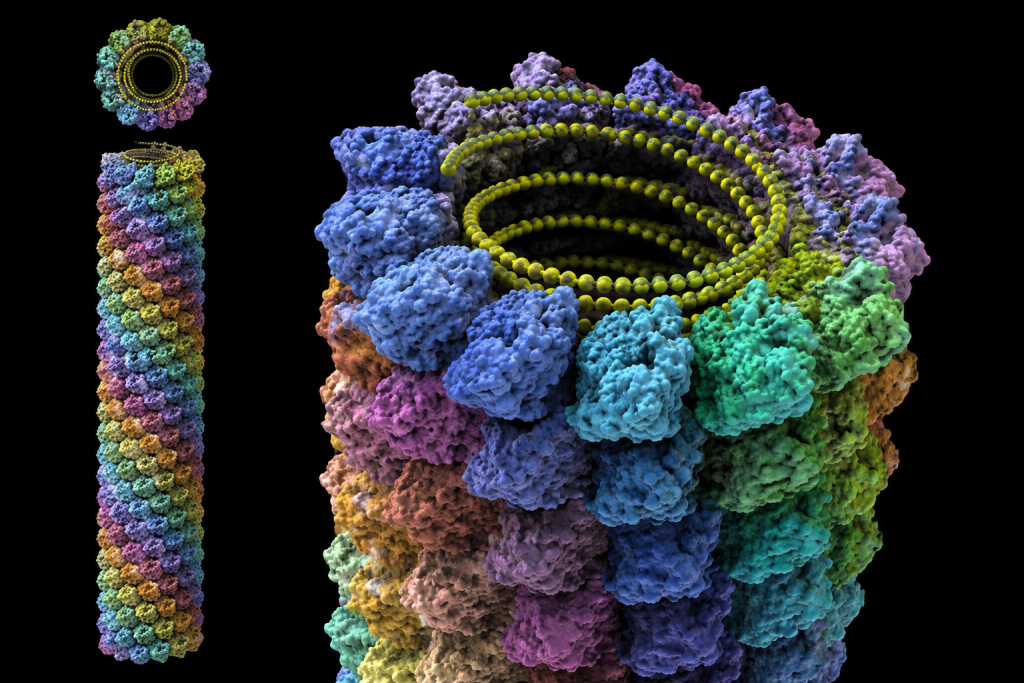
A classical example of a helical virus is the Virgaviridae, particularly Tobacco mosaic virus (TMV), which forms rigid, rod-shaped particles and is one of the most extensively studied viruses in molecular biology. TMV infects plants and causes characteristic mosaic-like discoloration of leaves, making it agriculturally significant.
Among animal viruses, members of the family Orthomyxoviridae, including Influenza A virus, possess helical nucleocapsids enclosed within an envelope. Although these viruses appear spherical externally due to their lipid envelope, the internal ribonucleoprotein structure maintains a helical arrangement.
Other important helical viruses include:
- Paramyxoviridae such as measles virus and mumps virus
- Rhabdoviridae such as rabies virus
- Filoviridae such as Ebola virus and Marburg virus
Helical symmetry represents an evolutionarily efficient structural strategy that enables viruses to package their genomes effectively while maintaining flexibility in shape, size, and host adaptation. Helical viruses possess several important biological and functional characteristics as follows:
- Flexible or filamentous morphology: Helical nucleocapsids often result in flexible, filamentous, or elongated virions. This flexibility is particularly evident in enveloped animal viruses, where the capsid structure adapts to the surrounding lipid membrane, allowing for pleomorphic shapes.
- Often enveloped in animal viruses: Most helical viruses that infect animals are enveloped, meaning they are surrounded by a lipid bilayer derived from the host cell membrane. This envelope contains viral glycoproteins that mediate attachment, fusion, and entry into host cells, enhancing infectivity but also increasing susceptibility to environmental degradation.
- Genome length directly influencing particle size: Because capsid proteins assemble along the nucleic acid strand, the length of the viral particle is directly determined by the length of the genome. This structural feature allows scalability without requiring major changes in capsid architecture.
- Efficient assembly due to repetitive protein subunits: Helical viruses benefit from highly efficient assembly mechanisms. Capsid proteins interact repetitively and sequentially with the genome, allowing rapid and energy-efficient formation of complete virions. This minimizes genetic complexity while ensuring reliable particle formation.
Complex viruses
Complex viruses represent a structurally diverse category of viruses that do not conform to the classical patterns of icosahedral or helical symmetry. Instead, they exhibit highly specialized and often elaborate architectures that combine multiple structural elements, reflecting advanced evolutionary adaptations to specific host environments and replication strategies. These viruses are particularly notable for their intricate organization, which may include multiple protein layers, accessory structures, and specialized infection apparatuses.
Unlike simpler virion forms, complex viruses possess structural components that cannot be described by a single geometric symmetry. Their morphology is often adapted to facilitate host recognition, genome delivery, and survival in challenging environments, particularly in bacterial and large DNA virus systems. Complex viruses demonstrate that viral architecture extends beyond simple geometric symmetry. Through highly specialized structural adaptations, these viruses achieve efficient genome delivery, autonomous replication (in some cases), and highly effective host targeting. Their diversity highlights the evolutionary flexibility of viral design and the intricate relationship between structure and function in virology.
Two of the most well-studied examples of complex viruses are members of the family Poxviridae and bacteriophages such as Myoviridae, especially bacteriophage T4, which infects bacterial cells.
Poxviruses (Family Poxviridae)
Members of the Poxviridae family are among the largest and most structurally complex animal viruses. They exhibit brick-shaped or ovoid virions with dimensions significantly larger than most other viruses. Their structure includes:
- A complex outer lipid membrane that encloses the virion
- Lateral bodies, which are protein-rich structures located between the core and the envelope
- A dense central core containing double-stranded DNA, which is the viral genome
The internal organization of poxviruses is highly specialized, allowing them to carry a large set of genes required for autonomous replication. Unlike most DNA viruses, poxviruses replicate entirely in the cytoplasm of the host cell, rather than relying on nuclear machinery. This unique capability is supported by the fact that they encode their own transcription enzymes, DNA-dependent RNA polymerases, and replication proteins, enabling them to perform independent gene expression outside the host nucleus. Examples include Variola virus (smallpox virus), Vaccinia virus, and Monkeypox virus (Mpox virus).
Bacteriophage T4 (Family Myoviridae)
Another classical example of a complex virus is bacteriophage T4, a member of the family Myoviridae, which infects Escherichia coli. Its structure is highly specialized for bacterial infection and consists of multiple distinct components:
- An icosahedral head that contains double-stranded DNA
- A contractile tail sheath that functions like a molecular syringe
- A base plate with tail fibers that recognize and bind bacterial surface receptors
Upon attachment to a bacterial cell, the tail sheath contracts, driving a hollow tube through the bacterial cell wall and membrane. This mechanism allows direct injection of viral DNA into the host cytoplasm, initiating infection with high efficiency. The structural complexity of T4 reflects its evolutionary adaptation for mechanical genome delivery in prokaryotic hosts.
Key characteristics of complex viruses
Complex viruses share several defining features:
- Non-uniform structural symmetry, combining multiple architectural elements
- Specialized infection machinery, such as tail structures or internal enzyme systems
- Large genome capacity, often encoding numerous accessory and replication-related proteins
- Independent replication capabilities in some cases, as seen in poxviruses
- Highly specialized host interaction mechanisms, particularly in bacteriophages
Bacteriophages: structural complexity and molecular infection machinery
Bacteriophages, or phages, are viruses that specifically infect bacteria. They represent some of the most structurally diverse and mechanically sophisticated viral entities. Their morphology is often classified as complex, as they do not conform strictly to simple icosahedral or helical symmetry. Instead, bacteriophages exhibit multi-component architectures that are highly adapted for efficient recognition, attachment, and genome delivery into bacterial cells. As aforementioned, one of the most extensively studied bacteriophages is Myoviridae, commonly referred to as bacteriophage T4. BacteriophageT4 infects Escherichia coli. Its structure is composed of several highly specialized functional regions that operate in a coordinated manner during infection (Figure 4).
Bacteriophages exemplify the highest level of structural specialization among viruses. Their modular architecture combining protective capsids, mechanical contractile systems, and receptor-specific appendages enables highly efficient infection of bacterial hosts. This complexity not only highlights viral evolutionary innovation but also provides valuable models for nanotechnology and molecular engineering research.
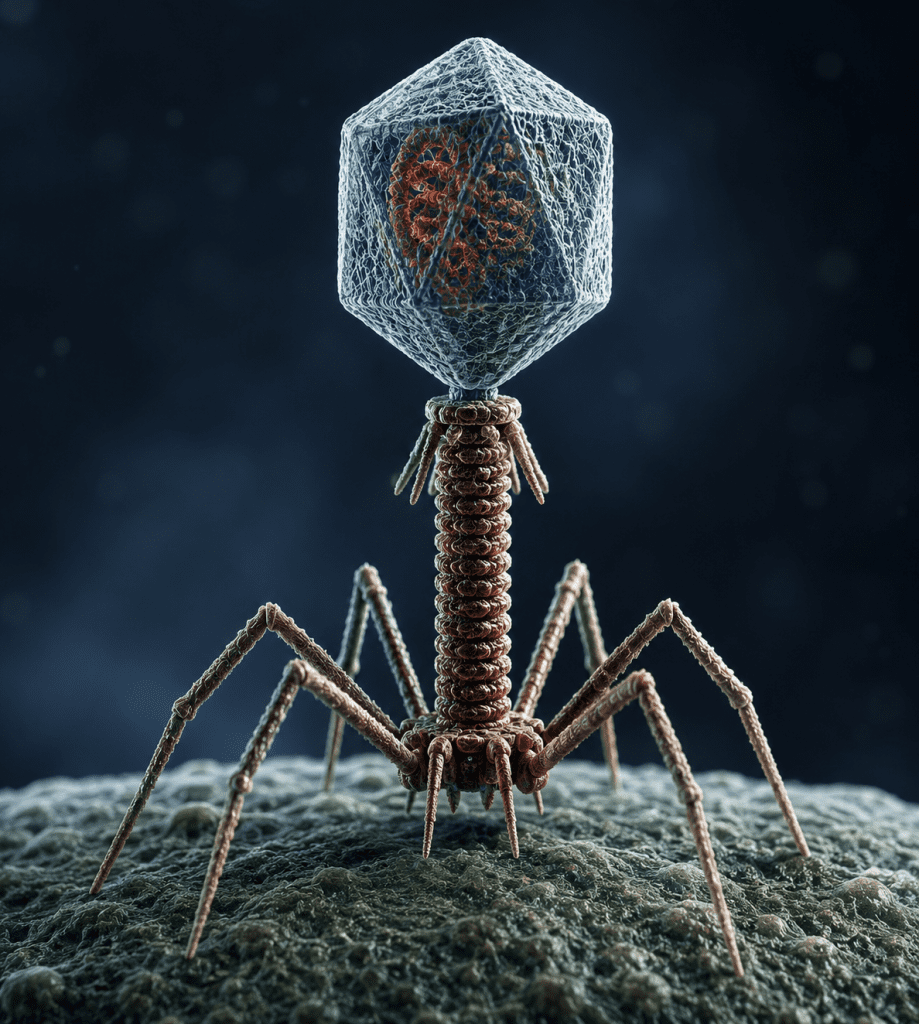
Figure 4. Structure of T4 bacteriophage, a complex virus. The different components of the phage are: baseplate, tail fibers, sheath, core, collar and head.
The morphology of bacteriophage T4 includes the following key structural components:
- Icosahedral head (capsid) containing DNA: The head region of bacteriophage T4 is an icosahedral capsid that encloses the double-stranded DNA genome. This structure provides a stable and protective environment for the viral genetic material, shielding it from degradation while ensuring efficient packaging at high density. The capsid is assembled from repeating protein subunits that self-organize into a robust polyhedral shell.
- Contractile tail sheath: Attached to the capsid is a long, cylindrical tail structure surrounded by a contractile sheath. This sheath functions as a dynamic mechanical component that undergoes rapid contraction during infection. When triggered, it shortens dramatically, driving internal components toward the bacterial surface.
- Tail fibers for host recognition and attachment: Extending from the base of the tail are tail fibers, which are responsible for recognizing and binding to specific receptors on the bacterial cell surface. These fibers determine host specificity and ensure that infection occurs only in suitable bacterial hosts. Their flexibility allows scanning of the bacterial surface for appropriate binding sites.
- Base plate for anchoring and activation: At the distal end of the tail lies the base plate, a complex structural platform that anchors the phage to the bacterial cell wall. It plays a critical role in initiating the infection process by stabilizing attachment and triggering tail sheath contraction.
Mechanism of infection of bacteriophage T4: molecular syringe action
During infection, bacteriophage T4 functions as a highly efficient molecular injection system. After attachment to the bacterial surface, structural changes in the base plate activate the contraction of the tail sheath. This contraction drives a central hollow tube through the bacterial cell wall and membrane, effectively puncturing the host cell envelope. Through this channel, the viral DNA is directly injected into the bacterial cytoplasm, initiating the infection cycle. This injection mechanism is often described as a molecular syringe, reflecting its precision, force generation, and efficiency in delivering genetic material into the host cell.
Enveloped viruses: structure, functional architecture, and biological implications
Enveloped viruses are a major structural class of viruses characterized by the presence of an outer lipid membrane known as the viral envelope. This envelope surrounds the underlying nucleocapsid and represents a critical adaptation that enhances viral infectivity, host-cell entry, and immune modulation. Unlike non-enveloped viruses, which rely solely on protein-based capsids for protection and transmission, enveloped viruses incorporate host-derived membrane components to form a biologically active interface with the environment and host cells.
The viral envelope is acquired during the process of viral budding, in which newly assembled virions exit the host cell by pushing through and encapsulating a portion of the host plasma membrane or internal membrane systems such as the endoplasmic reticulum or Golgi apparatus. As a result, the envelope is composed primarily of a phospholipid bilayer derived from the host cell, but it is extensively modified by viral components that are essential for infectivity.
Enveloped viruses represent a structurally sophisticated viral strategy in which a host-derived lipid membrane is repurposed to enhance infectivity, cellular entry, and immune interaction. While this envelope provides functional advantages in host infection and immune modulation, it also introduces structural fragility that limits environmental stability. The balance between enhanced infectivity and reduced resilience defines the biological and epidemiological behavior of enveloped viruses, making them a critical focus in virology, epidemiology, and antiviral research.
Structural components of the viral envelope
The envelope is not a passive covering; it is a highly functional and organized structure composed of three key elements:
1. Lipid bilayer: The foundational component of the envelope is a phospholipid bilayer, structurally similar to host cell membranes. This lipid layer provides flexibility and allows the virion to fuse with host cell membranes during infection. However, because it is derived from the host, it is also structurally fragile and sensitive to environmental conditions that disrupt lipid integrity.
2. Viral glycoproteins (spikes): Embedded within the lipid bilayer are viral glycoproteins, often referred to as spikes or peplomers. These are transmembrane proteins that extend outward from the viral surface and play a central role in host-virus interactions. They are among the most important determinants of viral infectivity and host specificity.
3. Matrix proteins: Located beneath the envelope is a layer of matrix proteins, which provide structural support and connect the nucleocapsid to the envelope. These proteins help stabilize virion assembly and coordinate the budding process by linking internal viral components with the external lipid membrane.
Functional roles of viral glycoproteins
Viral glycoproteins are essential for multiple stages of the viral life cycle and are among the most biologically active components of enveloped viruses:
- Host-cell recognition: Glycoproteins function as ligands that bind to specific receptors on host cell membranes. This receptor recognition determines host range and tissue tropism, ensuring that the virus infects only susceptible cells. The specificity of these interactions is a key determinant of viral pathogenicity and cross-species transmission.
- Membrane fusion: After attachment, viral glycoproteins mediate fusion between the viral envelope and host cell membrane. This fusion process allows the nucleocapsid or viral genome to enter the host cell cytoplasm. In some viruses, fusion occurs directly at the plasma membrane, while in others it occurs within endosomal compartments following endocytosis.
- Viral entry and internalization: Glycoproteins facilitate viral entry mechanisms, including receptor-mediated endocytosis and direct membrane fusion. These processes ensure efficient delivery of viral genetic material into the host cell, initiating replication.
- Immune system interaction: Glycoproteins are major antigenic determinants recognized by the host immune system. They stimulate antibody production and cellular immune responses. However, many enveloped viruses can alter or mask these proteins through mutation or glycosylation, enabling immune evasion and persistent infection.
Environmental sensitivity of enveloped viruses
A defining characteristic of enveloped viruses is their relative fragility outside the host organism. Because the envelope is composed of a lipid bilayer, it is highly susceptible to disruption by physical and chemical agents.
Enveloped viruses are particularly sensitive to:
- Heat, which destabilizes lipid structure and denatures envelope proteins
- Drying (desiccation), which disrupts membrane integrity
- Detergents and soaps, which dissolve lipid membranes
- Organic solvents (e.g., ether, alcohols), which extract membrane lipids
When the envelope is compromised, viral infectivity is lost because the glycoproteins required for attachment and fusion become non-functional and the nucleocapsid cannot effectively enter host cells. In contrast, non-enveloped viruses lack a lipid envelope and instead rely solely on their protein capsid for protection and host interaction. This structural simplicity provides significantly greater resistance to environmental stressors such as heat, acidity, drying, and chemical disinfectants. As a result, non-enveloped viruses often survive longer in external environments and may exhibit more efficient fecal-oral or contact-based transmission pathways.
Size and structural diversity of viruses
Viruses exhibit remarkable structural heterogeneity, particularly in terms of size and morphological organization. Despite being universally classified as acellular infectious agents, viral particles vary significantly in dimensions, ranging from extremely small virions to unusually large “giant viruses.” Most conventional viruses measure approximately 20 to 300 nanometers (nm) in diameter, placing them well below the resolution of light microscopy and necessitating electron microscopy for detailed visualization. However, certain giant viruses, such as members of the nucleocytoplasmic large DNA viruses (NCLDVs), can exceed 400 nm, approaching the size of small bacterial cells.
Viral size is not merely a physical attribute but a biologically significant factor that directly influences multiple stages of the viral life cycle. One major implication is the mode of entry into host cells. Smaller viruses often utilize receptor-mediated endocytosis or direct penetration mechanisms, whereas larger viruses may require more complex entry pathways or specialized structural adaptations.
Size also affects immune system detection and response. Smaller virions may evade early immune recognition due to limited antigenic surface exposure, while larger viruses present more extensive antigenic surfaces, potentially enhancing recognition by antibodies and immune effector cells. Viral size and structural diversity are fundamental determinants of infectivity, transmission efficiency, immune evasion, and disease manifestation, highlighting the close relationship between viral morphology and biological function.
Another important factor influenced by size is transmission dynamics. Smaller viruses are often more easily aerosolized and transmitted through respiratory droplets or airborne particles, whereas larger viruses may rely on direct contact or vector-mediated transmission. Additionally, viral size contributes to environmental persistence. In general, structural robustness, genome complexity, and capsid organization influence how long virions remain infectious outside a host organism.
Beyond size, structural diversity plays a critical role in viral biology. Variations in capsid symmetry (icosahedral, helical, or complex forms), presence or absence of an envelope, and organization of surface proteins all contribute to functional differences among viruses. Of particular importance are viral surface proteins, which determine receptor specificity on host cells. These interactions define host range and tissue tropism, ultimately influencing disease severity and pathogenic outcomes. For example, even closely related viruses may differ significantly in pathogenicity due to minor variations in surface glycoproteins that alter receptor binding affinity and immune recognition.
Functional significance of viral morphology
Viral morphology extends far beyond structural description and serves as a major determinant of viral infectivity, host interaction, transmission, pathogenicity, and clinical behavior. Features such as capsid symmetry, envelope composition, virion size, and surface protein organization directly influence how viruses recognize host cells, evade immune defenses, survive in external environments, and replicate within infected organisms. Consequently, understanding viral morphology is fundamental to virology, vaccine development, diagnostics, and disease control.
1. Host cell attachment and tropism
One of the most important functions of viral morphology is facilitating host-cell recognition and attachment. Viral surface structures including capsid proteins in non-enveloped viruses and glycoprotein spikes in enveloped viruses bind selectively to receptors located on host-cell membranes. This interaction determines host range (which species can be infected) and tissue tropism (which tissues or cell types are susceptible). Structural compatibility between viral attachment proteins and cellular receptors governs whether infection can occur. Even minor changes in viral surface architecture may alter receptor binding efficiency and influence transmissibility or disease severity.
2. Antigenicity and immune recognition
Viral structural proteins function as major antigenic determinants, meaning they are recognized by the host immune system and stimulate protective immune responses. Capsid proteins and envelope glycoproteins are common targets of neutralizing antibodies and cellular immunity. Because of their accessibility on the viral surface, these structural components form the basis of many modern vaccine strategies. Immune recognition of viral morphology therefore plays a central role in prevention, immunological memory, and long-term protection.
3. Environmental stability and survival
Viral morphology strongly influences survival outside the host. Non-enveloped viruses, protected by robust protein capsids, generally exhibit greater resistance to environmental stressors such as heat, drying, acidic conditions, and detergents. In contrast, enveloped viruses possess fragile lipid membranes that are easily disrupted, reducing environmental persistence but enhancing host-cell entry efficiency. These differences significantly affect patterns of transmission and epidemiology.
4. Replication strategy and genetic adaptation
Viral structure is closely associated with mechanisms of replication and genome expression. Certain structurally complex viruses possess additional functional capabilities that support independent replication processes. For example, bacteriophage T4, a structurally complex bacteriophage, contains specialized infection machinery and carries enzymes that facilitate efficient genome delivery and replication within bacterial hosts. Similarly, members of the family Orthomyxoviridae possess segmented RNA genomes, enabling genetic reassortment, a process that generates novel viral variants and contributes to rapid evolution.
5. Diagnostic identification and clinical applications
Morphological characteristics are also essential in laboratory diagnosis and viral identification. Techniques such as electron microscopy, immunofluorescence assays, and molecular diagnostic methods frequently rely on structural features to distinguish among viral groups. Capsid symmetry, envelope presence, virion dimensions, and antigenic profiles assist in classification and detection, supporting accurate diagnosis and surveillance. Viral morphology represents a direct link between structure and biological function, influencing virtually every stage of the viral life cycle and shaping both clinical outcomes and public health responses.
References
Acheson N.H (2011). Fundamentals of Molecular Virology. Second edition. John Wiley and Sons Limited, West Sussex, United Kingdom.
Alan J. Cann (2005). Principles of Molecular Virology. 4th edition. Elsevier Academic Press, Burlington, MA, USA.
Alberts B, Bray D, Johnson A, Lewis J, Raff M, Roberts K and Walter P (1998). Essential Cell Biology: An Introduction to the Molecular Biology of the Cell. Third edition. Garland Publishing Inc., New York.
Barrett J.T (1998). Microbiology and Immunology Concepts. Philadelphia, PA: Lippincott-Raven Publishers. USA.
Black, J.G. (2008). Microbiology: Principles and Explorations (7th ed.). Hoboken, NJ: J. Wiley & Sons.
Brian W.J Mahy and Mark H.C van Regenmortel (2010). Desk Encyclopedia of Human and Medical Virology. Elsevier Academic Press, San Diego, USA.
Brooks G.F., Butel J.S and Morse S.A (2004). Medical Microbiology, 23rd edition. McGraw Hill Publishers. USA.
Cann A.J (2011). Principles of Molecular Virology. Fifth edition. Academic Press, San Diego, United States.
Carter J and Saunders V (2013). Virology: Principles and Applications. Second edition. Wiley-Blackwell, New Jersey, United States.
Champoux J.J, Neidhardt F.C, Drew W.L and Plorde J.J (2004). Sherris Medical Microbiology: An Introduction to Infectious Diseases. 4th edition. McGraw Hill Companies Inc, USA.
Dimmock N (2015). Introduction to Modern Virology. Seventh edition. Wiley-Blackwell, New Jersey, United States.
Dimmock N.J, Easton A.J and Leppard K.N (2001). Introduction to modern virology. 5th edition. Blackwell Science publishers. Oxford, UK.
Discover more from Microbiology Class
Subscribe to get the latest posts sent to your email.