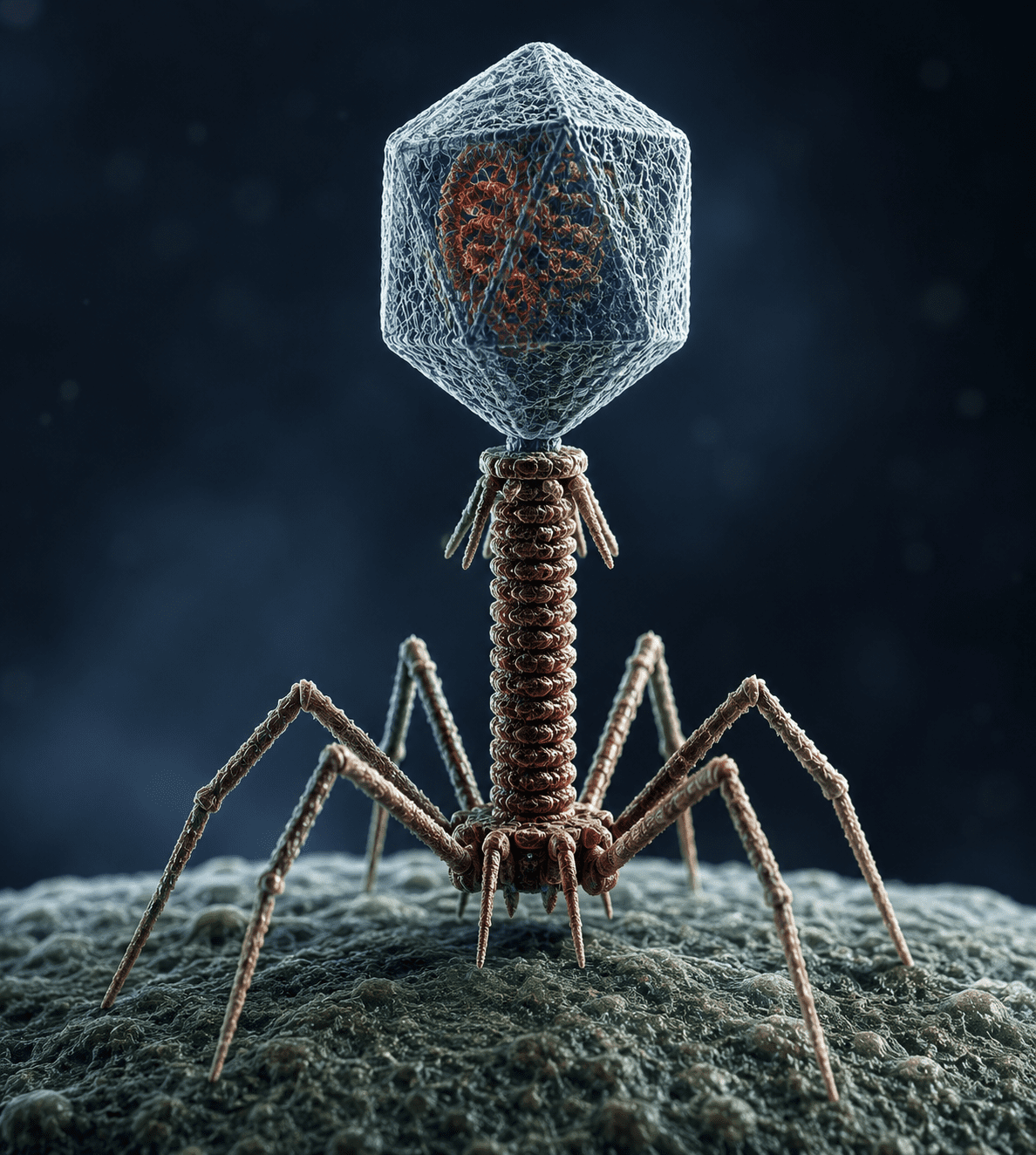
Viruses are acellular infectious agents that occupy a unique position at the interface of living and non-living biological systems. Their structural simplicity contrasts sharply with their biological impact, as they are capable of hijacking host cellular machinery to replicate and propagate. A virus is fundamentally composed of a nucleic acid genome, which may be either DNA or RNA, encased within a protective protein coat known as a capsid. In some cases, this core structure is further surrounded by a lipid envelope derived from host cell membranes, often studded with glycoproteins that mediate attachment and entry into susceptible cells.
The genetic material of viruses is highly diverse, both in composition and organization. This diversity extends to genome size, strand configuration, and replication strategies, allowing viruses to infect a broad spectrum of hosts ranging from bacteria to plants and animals. Unlike cellular organisms, viruses lack ribosomes, metabolic pathways, and the enzymatic systems required for independent energy production. As a result, they are entirely dependent on host cells for protein synthesis and metabolic intermediates, effectively rendering them obligate intracellular parasites.
The replication cycle of viruses is tightly coupled to host cell biology. After successful attachment to specific receptors on the host cell surface, viruses introduce their genetic material into the cell, where it is subsequently expressed and replicated using host-derived enzymes or virus-encoded proteins. Newly synthesized viral components are assembled into progeny virions, which are then released through mechanisms that may involve cell lysis or budding, depending on the virus type. This reliance on host systems not only ensures efficient replication but also often leads to cellular damage or altered physiological states in infected tissues.
Viruses exhibit a remarkable degree of specificity toward their hosts and even particular cell types within those hosts, a phenomenon largely dictated by molecular compatibility between viral surface proteins and host cell receptors. This specificity plays a central role in determining host range and tissue tropism, influencing patterns of infection and disease manifestation. Despite their simplicity, viruses demonstrate extensive evolutionary adaptability. High mutation rates, especially among RNA viruses, facilitate rapid genetic variation, enabling them to evade immune responses and develop resistance to antiviral agents. This evolutionary plasticity contributes to their persistence across diverse ecological niches and their continued relevance in both clinical and environmental contexts.
Ways in which viruses differ from other living cells
Viruses represent a fundamentally distinct biological entity when compared with cellular organisms such as bacteria, archaea, fungi, protozoa, and higher eukaryotic cells. While all cellular life forms share core organizational principles including autonomous metabolism, growth, and reproduction viruses lack many of these defining attributes. Instead, they function as obligate intracellular genetic elements that depend entirely on host cells for replication and survival. Their differences from living cells are structural, functional, and mechanistic, and these distinctions underpin their unique biological behavior.
The distinctions between viruses and living cells are profound and multifaceted, encompassing structural organization, metabolic capability, reproductive strategy, and interaction with host systems. While viruses exhibit certain life-like properties within host cells, their dependence on external cellular machinery and lack of autonomous biological functions place them in a unique category at the edge of classical definitions of life.
Dependence on living host cells for replication
One of the most defining characteristics that distinguishes viruses from all cellular organisms is their absolute dependence on living host cells for reproduction. Unlike bacteria or eukaryotic cells, which can replicate independently through binary fission or mitotic division, viruses are incapable of self-replication outside a host environment. They must first attach to a susceptible host cell, gain entry, and then exploit the host’s biosynthetic machinery to produce viral components. This obligate intracellular lifestyle makes them entirely dependent on cellular systems for transcription, translation, and energy supply. Without access to a living cell, viral particles remain inert and metabolically inactive.
Absence of cellular organization
Viruses are fundamentally acellular, meaning they do not possess the structural organization typical of living cells. They lack a cytoplasm, plasma membrane (in most cases, except for acquired envelopes), and internal organelles. Cellular organisms rely on compartmentalization for metabolic efficiency and regulation, but viruses are structurally simplified entities composed primarily of genetic material enclosed within a protein coat known as a capsid. Some viruses possess an additional lipid envelope derived from host membranes, but this structure does not function as an independent cellular boundary in the biological sense. The absence of cellular architecture places viruses outside traditional cell theory definitions of life.
Lack of metabolic machinery and energy production systems
Viruses do not possess metabolic pathways or enzymatic systems required for energy generation. Unlike living cells, which carry out respiration, fermentation, and other metabolic processes to produce ATP, viruses lack mitochondria, ribosomes, and enzymes necessary for independent biochemical activity. As a result, they are metabolically inert outside host cells. Once inside a host, they redirect cellular metabolism toward viral synthesis, effectively converting the host into a viral production unit. This reliance on host-derived energy and metabolites is a key distinction between viruses and living organisms.
Absence of ribosomes and protein synthesis capability
A critical feature that separates viruses from all cellular life forms is the absence of ribosomes. Ribosomes are essential molecular machines responsible for translating messenger RNA into proteins. Because viruses lack ribosomes, they cannot synthesize proteins independently. Instead, they rely entirely on host ribosomes to translate viral mRNA into structural and non-structural proteins. This dependence further reinforces their classification as obligate parasites and highlights their inability to carry out even the most basic functions of cellular life independently.
Unique genetic integration ability
Many viruses possess the ability to integrate their genetic material into the genome of their host cells. This process, known as integration, allows viral DNA or complementary DNA (in the case of RNA viruses via reverse transcription) to become part of the host’s chromosomal DNA. Once integrated, the viral genome may remain dormant for extended periods or be actively transcribed along with host genes. This phenomenon is particularly significant in the persistence of viral infections and in the alteration of host cellular function. Cellular organisms do not naturally integrate foreign genomes into their own as a standard part of their life cycle, making this a distinctive viral feature.
Distinct mode of reproduction
Viruses do not reproduce through conventional cellular division processes such as binary fission, mitosis, or meiosis. Instead, they follow a replicative cycle that involves disassembly, biosynthesis, assembly, and release. Viral components are synthesized separately within the host cell and later assembled into complete virions. This process is fundamentally different from cellular reproduction, which involves the duplication of all cellular components followed by division into two daughter cells. Viral replication is therefore a self-assembly process rather than a true reproductive event in the cellular sense.
Lack of independent metabolic system
In addition to lacking energy production mechanisms, viruses also do not maintain homeostatic regulatory systems. Cellular organisms regulate internal conditions such as ion balance, pH, and temperature through complex feedback mechanisms. Viruses, however, do not possess such regulatory capacities. Their structural and functional simplicity means that they cannot maintain internal equilibrium or respond independently to environmental changes. Instead, they remain dependent on the relatively stable internal environment of host cells.
Resistance to antibiotics and sensitivity to interferons
Viruses differ significantly from bacteria and other cellular pathogens in their response to antimicrobial agents. Antibiotics, which target bacterial structures such as cell walls, ribosomes, or metabolic pathways, are ineffective against viruses due to their lack of these structures. Instead, antiviral defense in hosts relies heavily on immune mechanisms, particularly interferons. Interferons are signaling proteins produced by host cells in response to viral infection. They function by inhibiting viral replication, enhancing immune recognition, and limiting viral spread within tissues. This distinction highlights the fundamentally different biological nature of viruses compared to antibiotic-sensitive microorganisms.
Extremely small size and ultramicroscopic nature
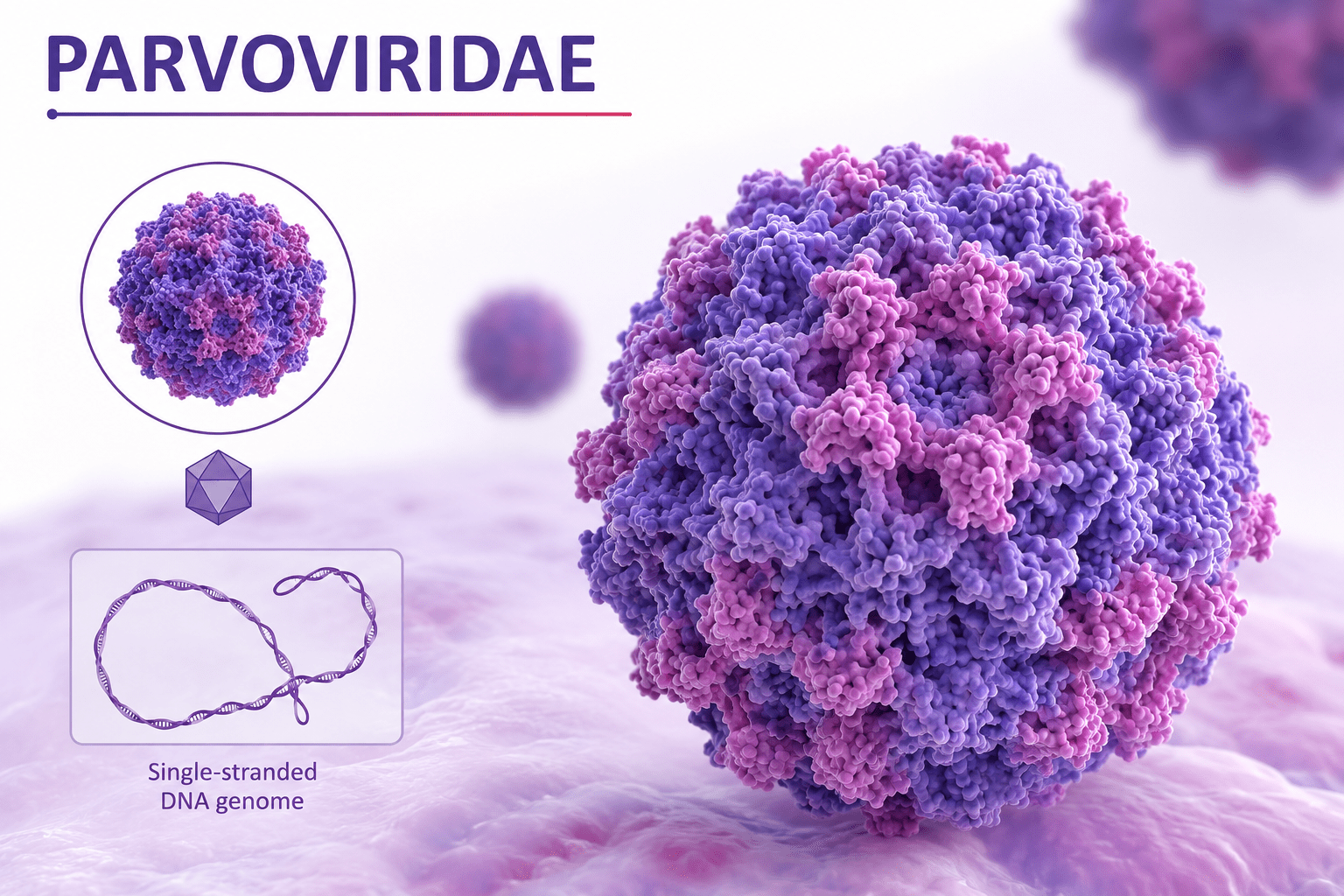
Viruses are among the smallest infectious agents known, typically ranging from approximately 20 to 300 nanometers in diameter. Their size places them well below the resolution limit of light microscopy, which is around 200-300 nanometers. Consequently, viruses can only be observed using electron microscopy, which provides the necessary resolution to visualize their structure. The small size of viruses is directly linked to their simplicity, as they contain only essential genetic and structural components required for infection and replication. Some of the smallest known viruses, such as parvoviruses, measure around 20 nanometers, while larger viruses such as poxviruses can reach up to 300 350 nanometers. The largest known viruses, such as mimiviruses (Figure 1), further blur the boundary between viral and cellular life due to their comparatively complex structure.
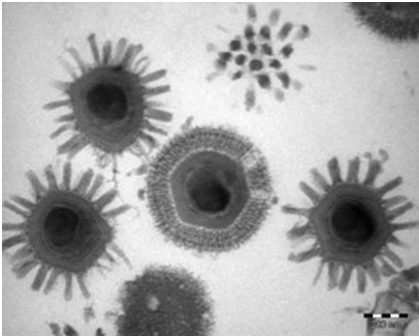
Genomic diversity and Baltimore classification
Viruses display an exceptional range of genomic diversity that is unmatched in cellular organisms. While all cellular life forms bacteria, archaea, and eukaryotes rely exclusively on double-stranded DNA (dsDNA) as their long-term genetic material, viruses are not constrained by a single nucleic acid type or replication strategy. Instead, they have evolved multiple genome forms that differ in chemical composition, strandness, polarity, and replication requirements. This diversity is not random but is functionally organized around how viral genetic material is converted into messenger RNA (mRNA), which is essential for protein synthesis.
To systematically describe this complexity, virologists use the Baltimore classification system, which groups viruses into seven mechanistic categories based on genome type and mRNA synthesis pathway.
- Double-stranded DNA viruses
- Single-stranded DNA viruses
- Double-stranded RNA viruses
- Positive-sense single-stranded RNA viruses
- Negative-sense single-stranded RNA viruses
- Reverse-transcribing RNA viruses
- Double-stranded DNA reverse-transcribing viruses
Double-stranded DNA (dsDNA) viruses
Double-stranded DNA viruses possess genomes composed of two complementary DNA strands, structurally resembling the genetic material of cellular organisms. This similarity allows them to exploit host nuclear machinery relatively efficiently. After entering a host cell, dsDNA viruses typically utilize host DNA-dependent RNA polymerases to transcribe viral genes into mRNA, although some larger dsDNA viruses encode their own replication enzymes.
Replication of dsDNA viruses generally occurs within the host nucleus, where viral DNA may exist either as an episome or become integrated into host chromosomal DNA in certain cases. These viruses can establish both lytic infections, where new virions are rapidly produced and released, and latent infections, where the viral genome persists with minimal gene expression. Examples include herpesviruses and adenoviruses. Their relatively stable genome structure allows for lower mutation rates compared to RNA viruses, contributing to long-term persistence in host populations.
Single-stranded DNA (ssDNA) viruses
Single-stranded DNA viruses have smaller and simpler genomes composed of a single DNA strand. Upon infection, ssDNA cannot be directly transcribed into mRNA. Therefore, it must first be converted into a double-stranded DNA intermediate using host DNA polymerases. Once this complementary strand is synthesized, normal transcriptional processes can proceed. This additional replication step introduces a dependency on host cell DNA synthesis machinery, often restricting ssDNA viruses to actively dividing cells. Parvoviruses are classic examples of this group. Despite their simplicity, ssDNA viruses are highly efficient in genome packaging and often have very small capsids, reflecting strong evolutionary pressure for minimal genetic content.
Double-stranded RNA (dsRNA) viruses
Double-stranded RNA viruses possess segmented or non-segmented genomes composed of two complementary RNA strands. Unlike DNA viruses, dsRNA cannot be directly translated or readily accessed by host ribosomes due to its structural stability and its tendency to trigger strong antiviral defenses.
To overcome this limitation, dsRNA viruses must carry their own RNA-dependent RNA polymerase within the virion. This enzyme synthesizes positive-sense mRNA from the dsRNA template immediately after infection. Without this pre-packaged enzyme, the viral genome would be biologically inactive inside the host. Reoviruses, including rotaviruses, are key representatives of this group and are often associated with gastrointestinal disease.
Positive-sense single-stranded RNA (+ssRNA) viruses
Positive-sense single-stranded RNA viruses possess genomes that function directly as messenger RNA. Upon entry into the host cell, their RNA can immediately bind to host ribosomes and initiate protein synthesis without prior transcription. This gives +ssRNA viruses a significant replication advantage, allowing rapid production of viral proteins.
Early translated proteins typically include RNA-dependent RNA polymerase, which is then used to synthesize complementary negative-strand RNA templates. These templates serve as intermediates for producing additional positive-sense genomes. This replication strategy is highly efficient and is characteristic of many medically important viruses such as coronaviruses and polioviruses. Because their replication is error-prone, +ssRNA viruses exhibit high mutation rates, which contributes to rapid evolution and frequent emergence of new strains.
Negative-sense single-stranded RNA (–ssRNA) viruses
Negative-sense single-stranded RNA viruses possess genomes that are complementary to mRNA and therefore cannot be directly translated. This makes them entirely dependent on viral machinery for initiating infection. As a result, these viruses must package an RNA-dependent RNA polymerase within the virion.
Once inside the host cell, this enzyme transcribes the negative-sense genome into positive-sense mRNA, which is then translated by host ribosomes. Viral replication proceeds through the synthesis of complementary RNA intermediates. Influenza viruses and rabies virus are prominent examples of this group. The requirement for virion-associated polymerase represents a key evolutionary adaptation that ensures immediate transcriptional capability upon infection.
Reverse-transcribing RNA viruses
Reverse-transcribing RNA viruses, primarily represented by retroviruses, possess RNA genomes that are not directly used as templates for protein synthesis. Instead, they carry the enzyme reverse transcriptase, which converts viral RNA into complementary DNA (cDNA) after infection. This newly synthesized DNA is then integrated into the host genome, forming a provirus. Once integrated, the viral DNA is transcribed by host RNA polymerase II, producing viral mRNA and genomic RNA. Human Immunodeficiency Virus (HIV) is the most well-known example of this group. This integration step allows long-term persistence in host cells and enables vertical transmission of viral genetic material during cell division, making these viruses particularly significant in chronic infections and oncogenesis.
Double-stranded DNA reverse-transcribing viruses
This group includes viruses that possess dsDNA genomes but replicate through an RNA intermediate. After infection, the viral DNA is transcribed into RNA, which is then reverse-transcribed back into DNA to produce new viral genomes. Hepatitis B virus is a classic example. This dual-phase replication strategy combines features of both DNA and RNA viruses and enhances genetic variability while maintaining genome stability.
Structural and functional implications of genomic diversity
Across all seven Baltimore groups, viral genomes vary not only in nucleic acid type but also in organization. Viral genomes may be linear or circular, segmented or non-segmented, and range from a few thousand nucleotides to over one million base pairs in giant viruses. Segmentation allows genetic reassortment, particularly in viruses like influenza, where co-infection can produce novel strains with altered virulence or host range. In contrast, cellular organisms maintain relatively uniform dsDNA genomes with stable replication and repair mechanisms. Viral genomic diversity therefore reflects adaptive evolutionary strategies rather than structural constraints of autonomous life.
Size, visualization, and structural efficiency
As aforesaid, viruses are among the smallest infectious agents known, typically ranging from approximately 20 to 300 nanometers. This places most viruses below the resolution limit of light microscopy, requiring electron microscopy for visualization. For example, parvoviruses are extremely small, while poxviruses are relatively large within the viral range. Some giant viruses, such as Mimivirus, challenge traditional size boundaries by approaching bacterial dimensions and possessing unusually large genomes. These discoveries have expanded the conceptual boundary between viruses and cellular organisms. Despite their simplicity, viral structures are highly efficient. Capsids are composed of repeating protein subunits arranged in symmetrical patterns such as icosahedral or helical forms, enabling structural stability with minimal genetic coding requirements. Enveloped viruses further acquire lipid membranes from host cells, incorporating glycoproteins essential for attachment and entry.
Host range and ecological significance
Viruses possess an extraordinarily broad host range and are capable of infecting virtually all known forms of life, including bacteria, archaea, fungi, plants, animals, and humans. This universal infective capacity distinguishes viruses from many other microorganisms, which are often restricted to narrower ecological niches. The ability of viruses to parasitize organisms across all biological domains demonstrates their remarkable adaptability and evolutionary success.
Among the most abundant and ecologically significant viruses are bacteriophages, or phages, which infect bacterial cells. Bacteriophages are considered the most numerous biological entities on Earth, with an estimated population exceeding 1031 particles globally. They are widely distributed in marine environments, freshwater systems, soils, and even within the microbiota of animals and humans. Phages play a crucial ecological role in regulating bacterial populations by lysing susceptible cells, thereby influencing nutrient cycling, microbial diversity, and ecosystem stability. In aquatic ecosystems, phage-mediated bacterial lysis contributes significantly to the release and recycling of organic matter, affecting global biogeochemical cycles such as carbon and nitrogen turnover.
Beyond population control, bacteriophages are major drivers of horizontal gene transfer among bacteria. Through processes such as transduction, phages transfer genetic material from one bacterial cell to another, facilitating the spread of genes associated with antibiotic resistance, virulence factors, and metabolic capabilities. This capacity has profound implications for microbial evolution and the emergence of pathogenic bacterial strains.
Viruses that infect plants are also of immense ecological and economic importance. Plant viruses interfere with normal physiological processes such as photosynthesis, nutrient transport, and growth regulation, leading to reduced crop yield and quality. Agricultural losses caused by plant viral diseases affect food security and global economies. Many plant viruses are transmitted by insect vectors such as aphids, whiteflies, and leafhoppers, creating complex interactions between viruses, hosts, and vector populations. Viral infections in plants may produce symptoms including chlorosis, mosaic patterns, leaf distortion, stunted growth, and tissue necrosis.
Animal and human viruses are responsible for a broad spectrum of infectious diseases ranging from mild, self-limiting illnesses to severe systemic and life-threatening conditions. Respiratory viruses, enteric viruses, neurotropic viruses, and hemorrhagic viruses all demonstrate the extensive pathological diversity associated with viral infections. Human viral diseases such as influenza, acquired immunodeficiency syndrome (AIDS), hepatitis, and coronavirus infections continue to pose major public health challenges worldwide.
The host range of a virus is largely determined by molecular compatibility between viral surface proteins and specific receptors on host cells. Viral attachment proteins must recognize and bind to complementary receptor molecules on the cell surface before entry can occur. This receptor specificity governs host susceptibility, tissue tropism, and disease manifestation. For example, some viruses exhibit narrow host ranges and infect only specific cell types within a single species, whereas others possess broader host ranges capable of crossing species barriers. Such cross-species transmission events, often referred to as zoonotic spillovers, are of particular significance because they can lead to the emergence of novel infectious diseases in human populations.
Evolutionary role and biological impact
Although viruses are structurally simple and lack independent cellular organization, they exert profound influence on biological evolution and genetic diversity. Viruses are not merely infectious agents; they are also important evolutionary forces that shape genomes, alter ecological relationships, and drive molecular innovation across living systems. One of the most significant evolutionary contributions of viruses is their role in horizontal gene transfer. During infection, viruses can acquire fragments of host genetic material and transfer them to new host cells in subsequent infections. This movement of genes between unrelated organisms accelerates genetic variation and facilitates the rapid spread of advantageous traits. In bacteria, viral-mediated gene transfer has contributed substantially to the dissemination of antibiotic resistance genes and virulence determinants.
Certain viruses also integrate their genomes into host chromosomal DNA, producing long-term genetic interactions between virus and host. In eukaryotic organisms, remnants of ancient viral infections are preserved within genomes as endogenous viral elements. In humans, endogenous retroviral sequences constitute a measurable fraction of the genome and provide evidence of historical viral integration events that occurred over millions of years. Some of these viral sequences have even acquired functional roles in host biology, illustrating how viral-host interactions can contribute to genomic innovation and evolutionary adaptation.
Viruses, particularly RNA viruses, are characterized by exceptionally high mutation rates due to the error-prone nature of viral polymerases. Unlike cellular DNA replication systems, many viral replication enzymes lack proofreading mechanisms, resulting in frequent genetic mutations during replication. This rapid accumulation of mutations enables viruses to adapt quickly to changing environmental conditions, evade host immune responses, and develop resistance to antiviral therapies.
The evolutionary flexibility of viruses contributes significantly to the emergence of new viral strains and variants. Processes such as mutation, recombination, and reassortment generate genetic diversity that may alter host range, virulence, or transmissibility. Influenza viruses, for instance, frequently undergo genetic reassortment, producing novel strains capable of causing epidemics or pandemics. Through their interactions with hosts and ecosystems, viruses influence biodiversity, population dynamics, and evolutionary trajectories across all domains of life. Their ability to reshape genomes, regulate microbial communities, and rapidly evolve ensures their continued significance in ecology, medicine, agriculture, and molecular biology.
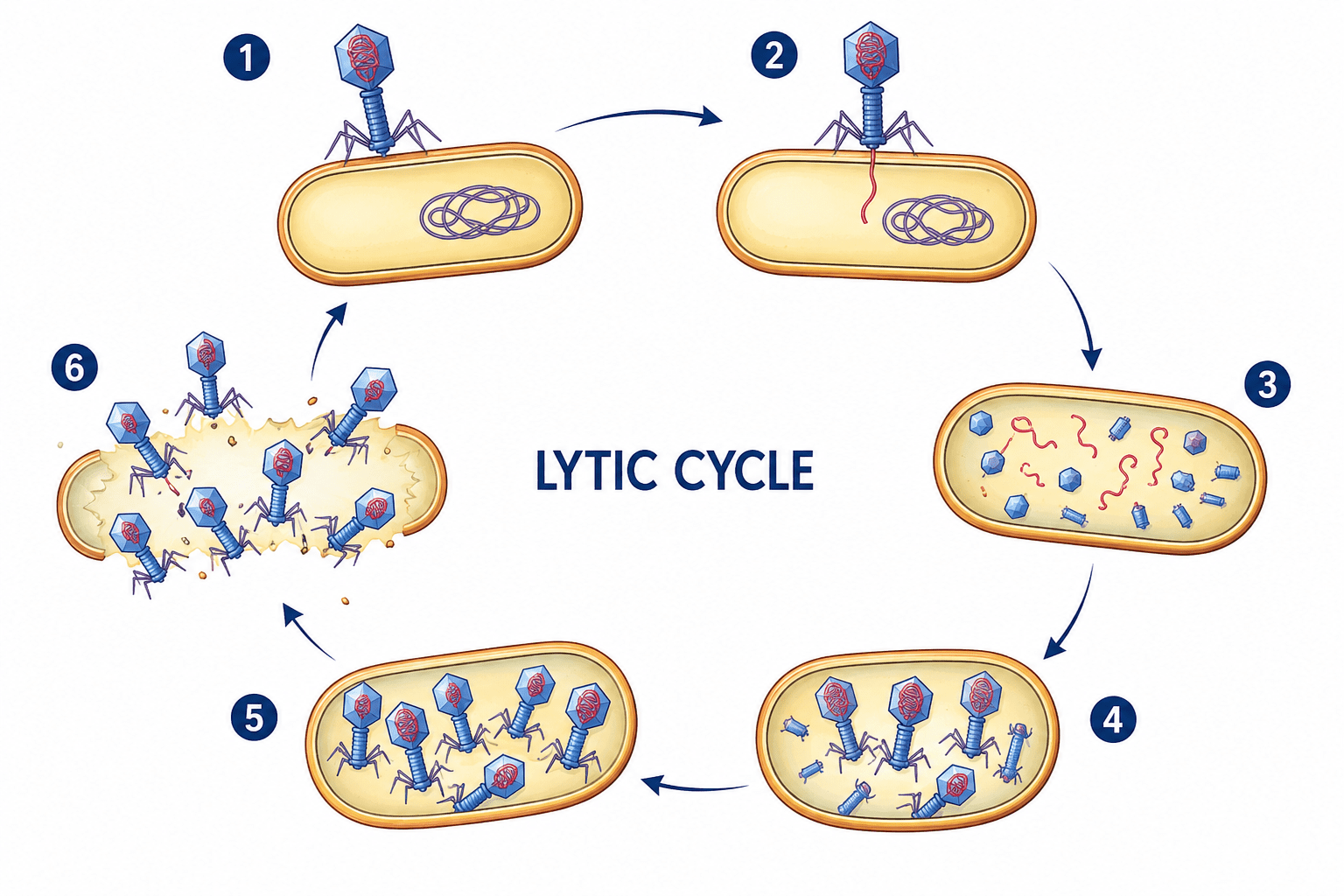
Viral replication and assembly process
Unlike cellular organisms, viruses do not reproduce through binary fission, mitosis, or any form of cellular division. Instead, viral multiplication occurs through a highly coordinated sequence of molecular events collectively referred to as the viral replication cycle. During this process, viruses exploit the biosynthetic and metabolic machinery of host cells to generate new infectious particles known as virions. Because viruses are acellular and metabolically inactive outside living cells, they cannot grow or divide independently. Rather than enlarging and splitting into daughter cells, new viral particles are synthesized component by component and subsequently assembled into complete virions. For this reason, viral multiplication is commonly described as a replicative assembly process rather than true cellular reproduction.
The viral replication cycle is a complex and highly coordinated process that differs fundamentally from cellular reproduction. Viruses do not grow or divide independently but instead utilize host cellular machinery to synthesize and assemble new virions from separate molecular components. Through sequential stages of attachment, penetration, uncoating, genome replication, protein synthesis, assembly, and release, viruses efficiently propagate within susceptible hosts despite their structural simplicity and metabolic dependence.
The different steps involved in viral replication cycles are:
- Attachment to a host cell
- Penetration and uncoating
- Genome replication
- Synthesis of viral proteins
- Assembly of new virions
- Release from the host cell
Attachment to the host cell
The first stage of viral replication is attachment, also known as adsorption. During this stage, viral surface proteins interact specifically with receptor molecules located on the surface of susceptible host cells. These receptors are usually normal cellular proteins, glycoproteins, or glycolipids that perform physiological functions for the host cell but are exploited by viruses as points of entry. The interaction between viral attachment proteins and host receptors is highly specific and determines the host range and tissue tropism of the virus. Only cells possessing compatible receptors can be infected successfully. For example, certain respiratory viruses specifically recognize receptors on epithelial cells of the respiratory tract, while neurotropic viruses target receptors on nerve cells. This receptor specificity explains why many viruses infect particular tissues or species selectively. Attachment is a critical determinant of infectivity because failure to bind effectively prevents subsequent stages of infection.
Penetration and uncoating
Following successful attachment, the virus gains entry into the host cell through a process known as penetration. The mechanism of entry varies depending on the type of virus and the nature of the host cell. Enveloped viruses commonly enter through membrane fusion, in which the viral envelope merges with the host cell membrane, allowing the viral nucleocapsid to enter the cytoplasm. Other viruses enter through receptor-mediated endocytosis, where the host cell engulfs the virus into intracellular vesicles. After penetration, the viral capsid is removed in a process called uncoating. During uncoating, viral nucleic acid is released from the protective protein coat into the host cell cytoplasm or nucleus. This step is essential because the viral genome must become accessible for transcription and replication. In some viruses, uncoating is mediated by host enzymes or acidic intracellular conditions that destabilize the capsid structure.
Genome replication
Once uncoated, the viral genome undergoes replication using either host or virus-encoded enzymes. The strategy employed depends largely on the type of viral nucleic acid. DNA viruses often replicate within the host nucleus using host DNA polymerases, whereas many RNA viruses replicate in the cytoplasm using viral RNA-dependent RNA polymerases. Positive-sense RNA viruses may use their genomes directly as messenger RNA, enabling immediate protein synthesis. Negative-sense RNA viruses must first synthesize complementary RNA strands before translation can occur. Retroviruses utilize reverse transcriptase to convert viral RNA into DNA, which then integrates into the host genome. Genome replication results in the production of numerous copies of viral nucleic acid that will later be packaged into newly formed virions.
Synthesis of viral proteins
In addition to genome replication, viruses direct the synthesis of viral proteins required for replication and assembly. Because viruses lack ribosomes and translational machinery, all viral proteins are synthesized by host ribosomes using viral messenger RNA. Early proteins synthesized during infection are often regulatory or enzymatic proteins involved in genome replication and suppression of host defenses. Later proteins generally include structural components such as capsid proteins, envelope glycoproteins, and matrix proteins necessary for virion formation. Many viruses alter host cellular metabolism extensively to prioritize viral protein production. In some cases, host protein synthesis is partially or completely inhibited, allowing the infected cell to function primarily as a viral production factory.
Assembly of new virions
Following synthesis of viral genomes and proteins, newly produced components are assembled into complete virions. Viral assembly is a highly organized process involving precise interactions between nucleic acids and structural proteins. Capsid proteins self-assemble into symmetrical structures, often exhibiting icosahedral or helical geometry. Viral genomes are selectively packaged into these capsids to form nucleocapsids. In enveloped viruses, nucleocapsids subsequently associate with modified host cell membranes containing viral glycoproteins. Unlike cellular organisms, viruses do not arise by division of pre-existing viral particles. Instead, every virion is constructed de novo from individually synthesized molecular components. This characteristic fundamentally distinguishes viral replication from cellular reproduction.
Release from the host cell
The final stage of the replication cycle is release of newly formed virions from the infected cell. Non-enveloped viruses are commonly released through cell lysis, a process in which the host cell ruptures and dies, liberating large numbers of viral particles. Enveloped viruses, however, are often released by budding through the host cell membrane. During budding, the virus acquires its lipid envelope from the host membrane while incorporating viral glycoproteins necessary for future infections. Budding may allow persistent infection without immediate destruction of the host cell. Once released, progeny virions are capable of infecting new susceptible cells, thereby perpetuating the infection cycle.
References
Acheson N.H (2011). Fundamentals of Molecular Virology. Second edition. John Wiley and Sons Limited, West Sussex, United Kingdom.
Alan J. Cann (2005). Principles of Molecular Virology. 4th edition. Elsevier Academic Press, Burlington, MA, USA.
Alberts B, Bray D, Johnson A, Lewis J, Raff M, Roberts Kand Walter P (1998). Essential Cell Biology: An Introduction to the Molecular Biology of the Cell. Third edition. Garland Publishing Inc., New York.
Barrett J.T (1998). Microbiology and Immunology Concepts. Philadelphia, PA: Lippincott-Raven Publishers. USA.
Black, J.G. (2008). Microbiology: Principles and Explorations (7th ed.). Hoboken, NJ: J. Wiley & Sons.
Brian W.J Mahy and Mark H.C van Regenmortel (2010). Desk Encyclopedia of Human and Medical Virology. Elsevier Academic Press, San Diego, USA.
Brooks G.F., Butel J.S and Morse S.A (2004). Medical Microbiology, 23rd edition. McGraw Hill Publishers. USA.
Cann A.J (2011). Principles of Molecular Virology. Fifth edition. Academic Press, San Diego, United States.
Carter J and Saunders V (2013). Virology: Principles and Applications. Second edition. Wiley-Blackwell, New Jersey, United States.
Champoux J.J, Neidhardt F.C, Drew W.L and Plorde J.J (2004). Sherris Medical Microbiology: An Introduction to Infectious Diseases. 4th edition. McGraw Hill Companies Inc, USA.
Dimmock N (2015). Introduction to Modern Virology. Seventh edition. Wiley-Blackwell, New Jersey, United States.
Dimmock N.J, Easton A.J and Leppard K.N (2001). Introduction to modern virology. 5th edition. Blackwell Science publishers. Oxford, UK.
Discover more from Microbiology Class
Subscribe to get the latest posts sent to your email.