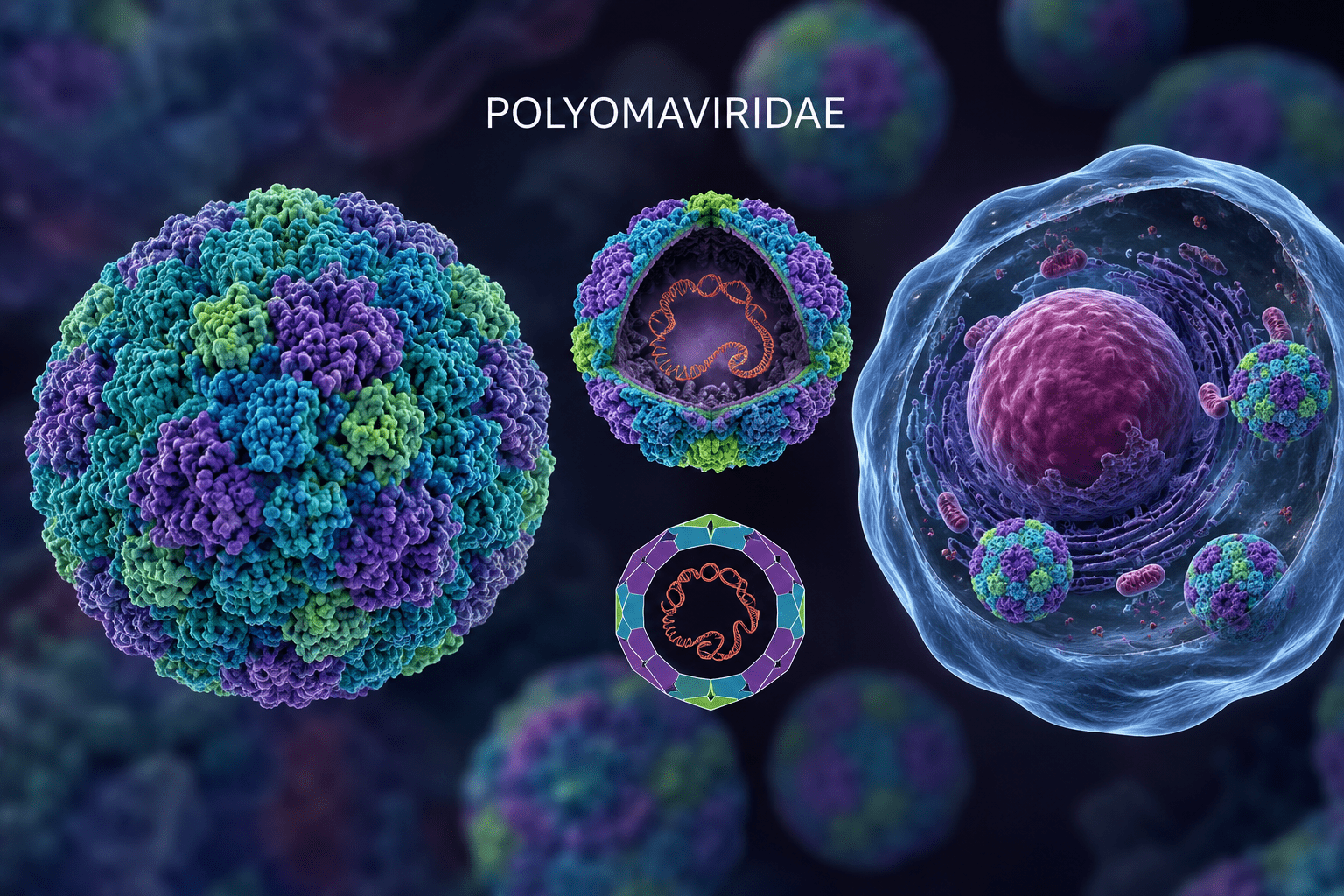
The family Polyomaviridae comprises a group of small, non-enveloped DNA viruses capable of infecting a wide range of vertebrate hosts, including humans, non-human primates, rodents, cattle, rabbits, and birds. Viruses in this viral family are known as polyomaviruses. Historically, viruses in the Polyomaviridae family attracted scientific interest because some members demonstrated the ability to induce tumours in experimental animals, giving rise to the term polyoma. The phrase ‘polyoma’ was derived from the Greek words poly (“many”) and oma (“tumours”). Although tumour formation is not a universal outcome of infection, the oncogenic potential of some polyomaviruses has remained an important feature of their biological significance.
Polyomaviridae was once classified together with papillomaviruses because both families contain small DNA tumour viruses. However, advances in molecular virology established them as distinct viral families with unique genomic organization, replication mechanisms, and host interactions. Members of Polyomaviridae are currently classified into multiple genera and species, with several known to infect humans. Human polyomaviruses include BK polyomavirus (BKPyV), JC polyomavirus (JCPyV), Merkel cell polyomavirus (MCPyV), and several recently identified species associated with specific tissues and disease states.
Polyomaviruses are widely distributed in nature and establish persistent or latent infections in their hosts. Most infections occur early in life and remain asymptomatic in immunocompetent individuals. Disease commonly develops when immune surveillance is impaired, allowing viral reactivation and tissue damage. Viruses in the Polyomaviridae family have gained increasing attention in transplant medicine, oncology, and studies of opportunistic viral infections.
Replication of polyomaviruses occurs primarily within the nucleus of infected host cells. Infection begins when viral capsid proteins bind specific receptors on the host-cell surface, followed by entry through receptor-mediated endocytosis. After internalization, the viral particle traffics through intracellular compartments until the viral DNA reaches the nucleus. Inside the nucleus, early genes are transcribed first using host-cell RNA polymerase. Production of T antigens initiates viral genome replication and modifies the host cellular environment to favour viral propagation. Following DNA replication, expression switches to late genes responsible for structural protein synthesis and virion assembly. Newly formed viral particles accumulate within the nucleus and are generally released through host-cell lysis, resulting in destruction of infected cells and dissemination of progeny virions.
Host specificity is an important biological feature of Polyomaviridae. Different polyomaviruses exhibit tissue tropism and species preferences. For example, BK polyomavirus primarily targets epithelial cells of the urinary tract and kidneys, whereas JC polyomavirus exhibits neurotropism and can infect glial cells of the central nervous system. Simian virus 40 (SV40), originally isolated from rhesus monkeys, became historically significant because of its accidental presence in some early poliovirus vaccine preparations produced in contaminated monkey kidney cell cultures. Although SV40 DNA has been detected in certain human samples, its role in human disease remains under scientific investigation.
The biology of the Polyomaviridae family reflects an efficient viral design characterized by compact genomic organization, nuclear replication, host adaptation, and the capacity for long-term persistence. These biological features continue to make polyomaviruses important models for understanding viral replication, oncogenesis, and host-virus interactions.
Structure of the Polyomaviridae family
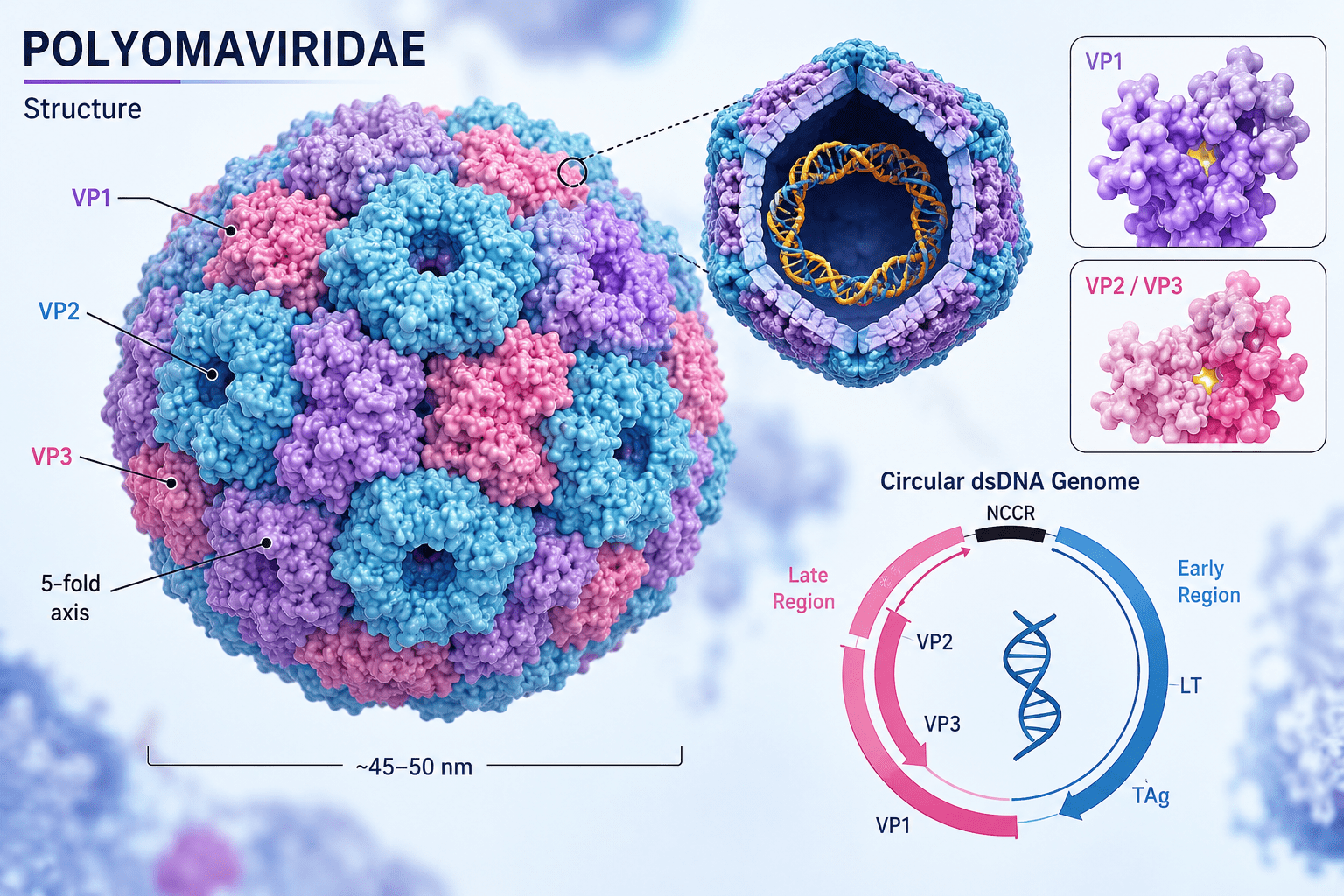
Members of the Polyomaviridae family are typically spherical and measure approximately 40-45 nm in diameter, making them among the smallest known animal viruses. Their compact structure is highly efficient and specialized for intracellular replication within the host nucleus. The virion consists primarily of a protein shell known as the capsid, which encloses and protects the viral genome. The capsid exhibits T=7 icosahedral symmetry and is composed of 72 capsomeres arranged in a highly organized geometric pattern. Because polyomaviruses lack a lipid envelope, the capsid forms the outermost layer of the virion and contributes significantly to environmental stability. As a result, these viruses are relatively resistant to ether, detergents, and adverse environmental conditions, although they remain susceptible to ultraviolet radiation, formalin, and other agents that damage nucleic acids or proteins.
The capsid is mainly constructed from the major structural protein VP1. Each capsomere consists of five VP1 molecules assembled into pentameric units, and interactions among these pentamers generate the stable icosahedral shell. VP1 is essential for maintaining capsid integrity and also mediates attachment of the virus to specific receptors on host cells. This receptor-binding activity plays a major role in determining host range, tissue tropism, and viral infectivity. Beneath the VP1 layer are two minor structural proteins, VP2 and VP3, which are present in smaller quantities. These proteins are internally located and contribute to virion assembly, genome packaging, and delivery of viral DNA into the host-cell nucleus during infection. Although less abundant than VP1, VP2 and VP3 are critical for successful viral entry and replication.
At the centre of the virion lies the viral genome (Figure 1), which consists of a circular double-stranded DNA (dsDNA) molecule approximately 5 kilobases in length. Despite its small size, the genome is highly organized and functionally efficient. It is divided into three principal regions: the early coding region, the late coding region, and the non-coding control region (NCCR). The early region encodes regulatory proteins known as tumour (T) antigens, principally the large T antigen and small T antigen. These proteins are synthesized early during infection and play central roles in viral replication and host-cell modulation. Large T antigen initiates viral DNA replication by binding to the viral origin of replication and recruiting host replication enzymes. It also alters host-cell cycle regulation, driving infected cells into a replication-competent state favourable for viral multiplication. Small T antigen enhances replication efficiency by modifying intracellular signalling pathways and cellular conditions that support viral growth.
The late region encodes the structural proteins VP1, VP2, and VP3 that form the viral capsid. Expression of these genes occurs later in the replication cycle, after sufficient viral DNA has been synthesized. This temporal regulation ensures coordinated assembly of mature virions and efficient packaging of newly replicated genomes. Separating the early and late regions is the non-coding control region, a highly important regulatory segment containing the origin of replication along with promoter and enhancer elements. The NCCR controls viral transcription and replication and strongly influences tissue tropism, pathogenicity, and replication efficiency. Genetic variations within this region may alter viral behaviour and contribute to differences in disease severity among polyomaviruses.
An unusual structural characteristic of polyomaviruses is the association of the viral DNA with host-derived histone proteins. Inside the virion, the circular dsDNA genome is organized into a chromatin-like complex resembling cellular chromosomes. This arrangement facilitates efficient interaction with host nuclear machinery and supports viral transcription, replication, and long-term persistence within infected cells. Polyomaviruses replicate within the nucleus of host cells, and their structural organization is well adapted for this intracellular environment. Following attachment and entry into the host cell, the capsid undergoes partial disassembly, allowing the viral DNA-histone complex to be transported into the nucleus. Viral replication and transcription then occur using host cellular enzymes and factors. Newly synthesized genomes are subsequently packaged into assembled capsids to form progeny virions, which are generally released through host-cell lysis. The structure of the family Polyomaviridae reflects a compact and highly efficient viral design. Their non-enveloped icosahedral capsid, chromatin-associated circular dsDNA genome, and specialized structural and regulatory proteins collectively enable successful infection, replication, persistence, and transmission across diverse vertebrate hosts.
Classification of human and non-human polyomaviruses
- BK polyomavirus (BKPyV): BKPyV is a ubiquitous human virus acquired in childhood and maintained as a latent infection, primarily in renal tissues. It becomes clinically important in immunosuppressed individuals, especially kidney transplant recipients, where it can cause polyomavirus-associated nephropathy and graft loss.
- JC polyomavirus (JCPyV): JCPyV is widely distributed globally and establishes lifelong latency in the kidneys, lymphoid tissue, and central nervous system. Reactivation in immunocompromised hosts can lead to progressive multifocal leukoencephalopathy (PML), a severe demyelinating brain disease.
- Merkel cell polyomavirus (MCPyV): MCPyV is a human skin-associated virus that integrates into host DNA and is strongly linked to Merkel cell carcinoma. Its oncogenicity is driven by viral T antigen expression following integration and mutation.
- SV40 (Simian virus 40): SV40 is a primate polyomavirus originally found in rhesus macaques. It gained attention due to historical contamination of early polio vaccines, and its role in human disease remains controversial.
- Trichodysplasia spinulosa-associated polyomavirus (TSPyV): TSPyV is associated with trichodysplasia spinulosa, a rare proliferative skin disorder occurring mainly in immunosuppressed patients. It persists latently and reactivates under immune suppression.
- KI polyomavirus (KIPyV): KIPyV is a respiratory polyomavirus detected in human airway samples worldwide. Its exact pathogenic role is unclear, although it is occasionally associated with respiratory illness.
- WU polyomavirus (WUPyV): WUPyV is another respiratory tract-associated virus discovered in human clinical specimens. It is frequently detected in children with respiratory symptoms, but causality remains uncertain.
- Human polyomavirus 6 (HPyV6): HPyV6 is a cutaneous polyomavirus commonly found on healthy human skin. It is generally non-pathogenic but may contribute to skin conditions in immunocompromised hosts.
- Human polyomavirus 7 (HPyV7): HPyV7 is also a skin-associated virus detected in the human cutaneous virome. Rare cases suggest a possible link to pruritic skin eruptions in immunosuppressed individuals.
- Human polyomavirus 9 (HPyV9): HPyV9 is detected at low levels in human blood and skin samples. Its clinical relevance remains unclear despite evidence of widespread exposure.
- Human polyomavirus 10 (HPyV10): HPyV10 is part of the diverse group of human skin-associated polyomaviruses. It has no confirmed disease association to date.
- Human polyomavirus 12 (HPyV12): HPyV12 has been identified in gastrointestinal and hepatic tissues. Its epidemiology and pathogenic potential are still under investigation.
- New Jersey polyomavirus: New Jersey polyomavirus was discovered in human stool samples and represents a genetically distinct lineage within the family. Its disease associations remain unknown.
- Lyon IARC polyomavirus: Lyon IARC polyomavirus was identified through molecular screening of human samples in research settings. No clear clinical disease has yet been attributed to it.
- Malawi polyomavirus: Malawi polyomavirus has been detected in respiratory and blood samples. Its epidemiological role and pathogenicity are still unclear.
- Saint Louis polyomavirus: Saint Louis polyomavirus is a newly identified human virus found in respiratory specimens. Its clinical significance has not been established.
- HPyV13: HPyV13 is a recently discovered human polyomavirus detected in limited samples. Its biological behavior and disease relevance remain largely unknown.
Pathogenesis of the Polyomaviridae family
The pathogenesis of the Polyomaviridae family is defined by asymptomatic primary infection, establishment of lifelong latency, immune-controlled persistence, and disease emergence following viral reactivation. Their capacity to manipulate host cell-cycle pathways, induce cell lysis, and exploit weakened immune defenses underlies the development of renal, urinary, neurological, and potentially oncogenic disease outcomes.
The Polyomaviridae family replicate within the nucleus of infected host cells. Although several members of this family infect a wide range of vertebrates, including mammals and birds, the human polyomaviruses particularly BK polyomavirus (BKPyV) and JC polyomavirus (JCPyV) are the most clinically significant members of the Polyomaviridae family because of their ability to establish persistent infections and cause severe disease under conditions of immune suppression. Pathogenesis within this viral family is characterized by viral persistence, cellular transformation potential, tissue-specific replication, and disease development associated primarily with reactivation rather than primary infection.
Polyomavirus infection generally begins through exposure during early childhood. Transmission routes are not completely understood but are believed to include respiratory, oral, urine-mediated, and possibly environmental exposure. Following entry into the host, the virus infects susceptible epithelial or lymphoid cells and establishes primary infection, which is often asymptomatic in immunocompetent individuals. Instead of causing immediate disease, the virus enters a latent or persistent state, allowing it to remain within the host for extended periods.
The pathogenesis of polyomaviruses depends heavily on their ability to establish lifelong persistence after initial infection. Once inside host cells, viral attachment occurs through interactions between viral capsid proteins and specific cellular receptors. Following receptor-mediated endocytosis, the virus enters the cell and traffics toward the nucleus where viral uncoating occurs. The circular dsDNA genome is then released into the nucleus and hijacks host cellular machinery for transcription and replication.
A central feature of polyomavirus pathogenesis is the expression of early regulatory proteins, particularly the large T antigen and small T antigen. These viral proteins play essential roles in driving host cells into active phases of the cell cycle to create conditions favorable for viral DNA replication. Large T antigen interacts with critical cellular regulatory proteins involved in growth control and DNA synthesis. By disrupting normal cell-cycle checkpoints, infected cells are induced to enter DNA synthesis phases, allowing viral replication to proceed efficiently.
This interference with cellular regulatory pathways explains why polyomaviruses are classified as DNA tumour viruses. Viral T antigens may interfere with tumour suppressor mechanisms and alter cellular signaling pathways, potentially leading to uncontrolled cellular proliferation. Although experimental studies have demonstrated oncogenic potential in animal models, the exact contribution of human polyomaviruses to human cancers remains an area of ongoing investigation.
Following genome replication and synthesis of structural proteins, newly assembled virions accumulate within the nucleus. Viral release occurs primarily through host cell lysis, leading to destruction of infected cells and subsequent tissue injury. The resulting pathological effects depend on the specific tissue tropism of the infecting polyomavirus and the immune status of the host.
For BK polyomavirus, pathogenesis primarily affects the urinary tract and kidneys. After initial infection, BKPyV establishes latency within renal tubular epithelial cells and urothelial tissues. In healthy individuals, viral replication remains controlled by cellular immunity and rarely causes disease. However, in immunocompromised states especially among transplant recipients receiving immunosuppressive therapy the virus can reactivate and undergo extensive replication.
Reactivation of BKPyV leads to direct cytopathic damage to renal epithelial cells. Progressive destruction of infected cells results in inflammation, tubular injury, and impaired renal function. This process may culminate in BK virus-associated nephropathy, characterized by declining kidney function and possible graft loss in renal transplant patients. BKPyV may also infect the bladder epithelium, producing hemorrhagic cystitis characterized by inflammation, epithelial destruction, and bleeding.
In contrast, JC polyomavirus exhibits neurotropism and preferentially targets the central nervous system during reactivation. JCPyV remains latent in sites such as the kidneys, bone marrow, and lymphoid tissues following primary infection. In individuals with profound immune suppression including patients with advanced HIV infection, hematological malignancies, or immunomodulatory therapy the virus may reactivate and spread to the brain.
Within the central nervous system, JCPyV infects oligodendrocytes, the cells responsible for producing myelin. Viral replication causes lytic destruction of these cells, resulting in progressive demyelination. This pathological process leads to progressive multifocal leukoencephalopathy (PML), a severe neurological disorder characterized by motor dysfunction, cognitive decline, visual impairment, speech disturbances, and progressive neurological deterioration. Because oligodendrocyte loss is irreversible and viral replication may continue unchecked in immunosuppressed patients, PML is associated with high morbidity and mortality.
Another notable member of this family is Simian Virus 40 (SV40), originally identified in rhesus monkeys. SV40 gained attention because of historical contamination of early poliovirus vaccines produced in infected monkey kidney cell cultures. SV40 demonstrates strong transforming capabilities in experimental systems due to the activity of its T antigens. Viral proteins can interfere with cellular growth regulation and promote immortalization of infected cells. Although SV40 DNA has been detected in some human biological samples, its direct pathogenic significance in humans remains controversial and has not been conclusively established.
Detection of Polyomaviridae infections
Accurate detection of Polyomaviridae infections relies on a combination of molecular, virological, and histopathological approaches, as these viruses often establish persistent or latent infections that are difficult to identify through routine clinical screening. The most clinically significant human members include BK polyomavirus (BKPyV), JC polyomavirus (JCPyV), and the less clearly pathogenic SV40-like polyomaviruses. A central diagnostic challenge is that reactivation typically occurs in immunocompromised individuals, where viral replication becomes detectable in bodily fluids or tissues.
The primary diagnostic modality is nucleic acid amplification testing (NAAT), particularly polymerase chain reaction (PCR), which is widely used to detect viral DNA in urine, blood (plasma or serum), cerebrospinal fluid (CSF), and tissue biopsies. For BK polyomavirus, quantitative PCR of plasma and urine is the standard for monitoring viral load, especially in renal transplant recipients where BK-associated nephropathy is a major complication. Rising viral loads in plasma correlate strongly with progressive renal dysfunction and are used as an early predictive biomarker before irreversible kidney damage occurs.
For JC polyomavirus, detection is more specialized due to its neurotropism. CSF PCR is the gold standard for diagnosing progressive multifocal leukoencephalopathy (PML). However, sensitivity may vary depending on disease stage and viral burden in the central nervous system. In cases where PCR is negative but clinical suspicion remains high, brain biopsy with histopathology may be required. Characteristic findings include demyelination, enlarged oligodendrocyte nuclei with viral inclusions, and atypical astrocytes. Immunohistochemistry using antibodies against SV40 large T-antigen is often employed as a cross-reactive marker for polyomavirus infection.
Serological testing plays a limited but supportive role. While most adults have antibodies due to prior exposure, serology cannot reliably distinguish between latent and active infection. Instead, it is more useful for epidemiological studies than for clinical decision-making. In transplant medicine, routine surveillance using PCR-based viral load monitoring is preferred over serology.
Advanced techniques such as next-generation sequencing (NGS) are increasingly used in research and complex diagnostic cases, particularly when atypical presentations occur or when multiple opportunistic infections are suspected. NGS can detect low-level viral DNA and identify co-infections that may influence disease progression. Detection strategies for polyomaviruses are centered on quantitative molecular diagnostics combined with tissue-based confirmation, with emphasis on viral load dynamics as a marker of disease activity rather than simple presence or absence of infection.
Treatment and clinical management of Polyomaviridae infections
The treatment of Polyomaviridae infections is fundamentally immune-centered rather than virus-targeted, relying on restoration of host antiviral immunity, cautious modulation of immunosuppression, and selective use of adjunctive antivirals with limited efficacy. There are currently no virus-specific antiviral drugs that reliably eradicate Polyomaviridae infections, and management is largely based on controlling viral replication through immune restoration and supportive care. Treatment strategies differ depending on the infecting virus, affected organ system, and immune status of the host.
For BK polyomavirus-associated nephropathy (BKVAN), which is most commonly seen in kidney transplant recipients, the primary therapeutic approach is reduction of immunosuppression. Since viral reactivation is driven by immune suppression, careful tapering of agents such as calcineurin inhibitors or antimetabolites can restore immune control and reduce viral replication. This must be balanced against the risk of graft rejection, making management highly individualized.
Adjunctive therapies have been explored but with variable success. Leflunomide, an immunomodulatory drug with antiviral properties, has been used off-label in some cases. Cidofovir, a nucleotide analogue with broad antiviral activity, has also been employed, although nephrotoxicity limits its use, particularly in patients with existing renal impairment. Intravenous immunoglobulin (IVIG) is sometimes administered to provide passive immunity, though evidence for consistent benefit remains limited.
For JC polyomavirus infection leading to progressive multifocal leukoencephalopathy (PML), treatment is even more challenging. The cornerstone of therapy is immune reconstitution, particularly in HIV-associated cases where initiation or optimization of highly active antiretroviral therapy (HAART) can lead to partial immune recovery and improved survival. In non-HIV immunosuppressed patients, reversal or reduction of immunosuppressive therapy is similarly critical.
Despite immune recovery being the most effective intervention, neurological damage from PML is often irreversible, and mortality remains high. Experimental therapies such as checkpoint inhibitors (e.g., PD-1 blockade) have been investigated to enhance antiviral T-cell responses, with some promising case reports but no definitive standard of care established. Supportive care remains essential across all polyomavirus infections. This includes renal function monitoring in BK infection, neurological and radiological surveillance in PML, and management of secondary complications such as opportunistic infections. Rehabilitation services may also be required for survivors of CNS involvement.
Prevention and control of Polyomaviridae
Prevention and control of viruses in the family Polyomaviridae are important because several members, such as BK virus, JC virus, and Merkel cell polyomavirus, can cause severe disease in immunocompromised individuals. Since these viruses are widespread in human populations and often remain latent after primary infection, complete eradication is difficult. Therefore, control strategies mainly focus on limiting transmission, reducing viral reactivation, and protecting vulnerable patients.
Good hygiene practices are essential for prevention. Frequent hand washing, proper sanitation, and disinfection of contaminated surfaces help reduce viral spread through urine, respiratory secretions, or direct contact. In healthcare settings, strict infection-control measures, including sterilization of medical equipment and isolation precautions when necessary, can minimize transmission among patients.
Monitoring immunocompromised individuals is a major control strategy. Organ transplant recipients, cancer patients, and people with weakened immune systems are at higher risk of polyomavirus-associated diseases. Regular screening for viral DNA using molecular diagnostic techniques such as PCR allows early detection of viral reactivation. Early diagnosis helps clinicians prevent severe complications such as BK virus nephropathy or progressive multifocal leukoencephalopathy (PML) caused by JC virus.
Management of immunosuppression is also critical. Reducing immunosuppressive therapy, when clinically possible, may help restore immune responses and limit viral replication. Currently, no highly effective antiviral drug specifically targets polyomaviruses, although supportive therapies and experimental antiviral agents are sometimes used.
Research into vaccines and targeted antiviral therapies is ongoing. Advances in molecular virology and immunotherapy may improve future prevention and treatment approaches. Public health education, early diagnosis, routine surveillance in high-risk populations, and strong hospital infection-control programs remain the most effective methods for controlling diseases associated with Polyomaviridae.
References
Acheson N.H (2011). Fundamentals of Molecular Virology. Second edition. John Wiley and Sons Limited, West Sussex, United Kingdom.
Alan J. Cann (2005). Principles of Molecular Virology. 4th edition. Elsevier Academic Press, Burlington, MA, USA.
Alberts B, Bray D, Johnson A, Lewis J, Raff M, Roberts K and Walter P (1998). Essential Cell Biology: An Introduction to the Molecular Biology of the Cell. Third edition. Garland Publishing Inc., New York.
Barrett J.T (1998). Microbiology and Immunology Concepts. Philadelphia, PA: Lippincott-Raven Publishers. USA.
Black, J.G. (2008). Microbiology: Principles and Explorations (7th ed.). Hoboken, NJ: J. Wiley & Sons.
Brian W.J Mahy and Mark H.C van Regenmortel (2010). Desk Encyclopedia of Human and Medical Virology. Elsevier Academic Press, San Diego, USA.
Brooks G.F., Butel J.S and Morse S.A (2004). Medical Microbiology, 23rd edition. McGraw Hill Publishers. USA.
Cann A.J (2011). Principles of Molecular Virology. Fifth edition. Academic Press, San Diego, United States.
Carter J and Saunders V (2013). Virology: Principles and Applications. Second edition. Wiley-Blackwell, New Jersey, United States.
Champoux J.J, Neidhardt F.C, Drew W.L and Plorde J.J (2004). Sherris Medical Microbiology: An Introduction to Infectious Diseases. 4th edition. McGraw Hill Companies Inc, USA.
Dimmock N (2015). Introduction to Modern Virology. Seventh edition. Wiley-Blackwell, New Jersey, United States.
Dimmock N.J, Easton A.J and Leppard K.N (2001). Introduction to modern virology. 5th edition. Blackwell Science publishers. Oxford, UK.
Discover more from Microbiology Class
Subscribe to get the latest posts sent to your email.